hemoglobina
Hemoglobina (Hb) este pigmentul transportor de oxigen in
eritrocitele vertebratelor, fiind
elementul esential pentru principala functie fiziologica a
acestora, functia respiratorie. Reprezinta 95% din proteinele
solubile ale eritrocitului si 35% din greutatea acestuia, astfel incat se
poate spune ca eritrocitul este format doar dintr-o membrana, care
protejeaza o solutie concentrata de Hb. Localizarea in
eritrocite prezinta doua avantaje:
- un transport eficient al O2
fara cresterea presiunii osmotice a plasmei asa cum s-ar
intampla daca Hb ar fi o simpla proteina plasmatica;
- in eritrocite exista un mecanism de
reducere a Fe3+ din methemoglobina la Fe2+, acest
mecanism fiind reprezentat de NADH-methemoglobin reductaza.
Valori normale: ♂ 13 - 17g/dl; ♀ 13 - 16g/dl. Un
barbat de 70 Kg are aprox. 900g de Hb. Aceste valori corespund la o :
- valoare medie pe hematie - HEM = 29 mmg;
- valoare medie/100 ml masa
eritrocitara - CHEM=34%
Structura chimica si sinteza
HEMOGLOBINEI
Hb este o hemoproteina, cu GM=64000
daltoni. Molecula de Hb are forma globulara si este formata
dintr-o componenta proteica-globina
(96% din molecula) si o metaloporfirina-hemul, care contine Fe2+ (feros) de culoare
rosie. Componentele necesare sintezei de Hb sunt: globina, protoporfirina
IX, fierul si 2,3 difosfogliceratul (2,3-DPG).
Hemul. Din punct de vedere chimic este format dintr-un atom de Fe2+
fixat in centrul unui nucleu tetrapirolic, acelasi ca pentru
clorofila si vit. B12.
Patrulaterul, in centrul caruia se afla Fe2+ are in
unghiuri cei 4 atomi de N ai nucleilor pirolici (figura 5.1.). Generarea
hemului necesita, ca pas esential, formarea protoporfirinei IX,
ultimul compus al unui lung sir de reactii enzimatice, care se
desfasoara alternativ in mitocondriile eritroblastilor
nucleati si
in citoplasma celulara. Sinteza incepe in stadiul de eritroblast bazofil,
este maxima in eritroblastii policromatofili si oxifili, foarte
redusa in reticulocite si absenta in eritrocitul matur, care nu
mai are mitocondrii si isi pierde o parte din echipamentul enzimatic
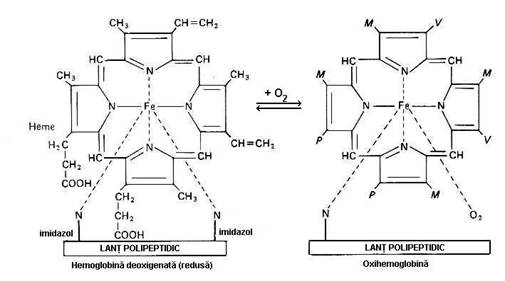
Figura 5.1. Formele spatiale ale hemoglobinei.
Abrevierile M, V si P reprezinta radicalii moleculei din stanga (dupa Ganong, F. W, 1993).
Hemul format este expulzat din mitocondrii in
citopasma, unde 4 grupari hem se leaga de 4 lanturi de
globina pentru a forma molecula de Hb.
Structura hemului este constanta in
diversele tipuri de Hb, necunoscandu-se variante structurale conditionate
genetic ca la globina.
Datorita proprietatii sale
de a forma legaturi reversibile cu O2, hemul reprezinta
componenta fiziologic activa din molecula de Hb.
Hemul
se mai gaseste si in structura: mioglobinei-pigmentul purtator de O2 din muschii
rosii si in enzimele lantului respirator-citocromul c.
Sinteza hemului este autocontrolata prin
feed-back negativ (cresterea concentratiei hemului determina
depresia enzimei ALA-sintetaza si productia de hem
inceteaza.
Sinteza defectuasa a hemului se
exprima sub forma de porfirii.
Porfiriile sunt
maladii datorate deficientei ereditare, congenitale sau dobandite a
enzimelor care intervin in sinteza hemului si se caracterizeaza prin
acumularea de precursori porfirinici sau unul sau mai multi porfirinogeni
in maduva hematogena (porfiria eritropoietica) sau in ficat (porfiria
hepatica), consecinta fiind scaderea productiei de Hb.
Simptomul dominant este reprezentat de leziunile cutanate fotosensibile, la
care se pot adauga semne de anemie sau suferinta hepatica
Globina este o proteina cu caracter bazic
din subclasa histonelor. Se sintetizeaza la nivel ribozomal, sinteza fiind
controlata de acizii nucleici pe caile obisnuite ale sintezei
proteinelor.
Globina este diferita ca
structura de la un tip de Hb la alta, fiind componenta care imprima specificitatea diverselor Hb cunoscute. Diferenta de
structura nu consta in numarul lanturilor, toate globinele
sunt tetrameri (au 4 lanturi polipeptidice, identice doua cate
doua), ci in numarul si succesiunea aminoacizilor, adica in
structura primara a moleculei de globina. Lanturile
polipeptidice sunt desemnate prin literele grecesti α, β,
γ, δ, ε, si ζ, principalele deosebiri dintre aceste
lanturi fiind redate in tabelul 5.1.
|
Denumirea
greceasca
|
Numele grecesc
|
Numarul de aminoacizi
|
Comentarii
|
|
Alfa
|
|
| |
|
Beta
|
|
| |
|
Gamma
|
|
Difera de
lanturile β prin 39 de aminoacizi
| |
|
Delta
|
|
Difera de
lanturile β prin 10
aminoacizi
| |
|
Epsilon
|
|
Numai in viata
embrionara (primele 3 luni)
| |
|
Zeta
|
|
Numai in viata
embrionara (primele 3 luni)
| |
|
|
|
|
|
| | | | | | | | |
Tabelul nr. 5.1. Lanturile polipeptidice prezente in diferitele tipuri
de Hb.
Fiecare molecula
de Hb are 2 perechi de polipeptide neidentice (lanturi):
- 2 lanturi a, a caror
sinteza este dublu codificata de 2 gene identice aflate pe cromozomul
16, fiecare lant a avand 141 de aminoacizi;
- 2 lanturi
non- a, care
pot fi β, γ, δ, ε, si ζ a caror sinteza
este codificata de gene aflate pe cromozomul 11; fiecare din aceste
lanturi poseda 146 de aminoacizi, dar in succesiune diferita.
Dupa sinteza la nivel ribozomal,
lanturile libere sunt incorporate in molecula completa de Hb rapid
si spontan, nefiind necesara prezenta altor factori. Fiecare
subunitate polipeptidica este legata de un hem, deci o molecula
de Hb contine 4 grupari hem, fiecare grupare hem avand in centru un
atom de Fe2+ (in total 4 atomi de Fe2+).
Structura
primara a Hb-reprezinta secventa
aminoacizilor in structura fiecarui lant polipeptidic si legarea
Fe2+ de histidina acestui lant.
Structura
secundara (spiralele lui Pauling si Corey)
este reprezentata de configuratia spatiala,
helicoidala a fiecarui lant globinic. Pe o lungime de
aproximativ 75%, lanturile de aminoacizi au o structura
spatiala de tip a-helicoidal, care alterneaza la anumite intervale cu segmente
nehelicoidale, la nivelul carora se produce indoirea si
incolacirea catenei.
Structura
tertiara este reprezentata de pliurile
(buzunarele hemului) care adapostesc gruparile hem ale fiecarui
lant globinic.
Structura
cuaternara rezulta din aranjarea celor 4
monomeri aflati in molecula de Hb, astfel incat formeaza o
structura globulara compacta. Aceasta prezinta in
apropierea suprafetei extreme 4 cavitati numite pliurile sau
pungile hemului, care adapostesc cate un hem. In lungul axei moleculei de
Hb, separand lanturile unul de celalalt se gaseste o
cavitate centrala in care se fixeaza 2,3 DPG. Structura tridimensionala
a Hb a fost determinata prin analiza cristalografica cu raze X
(figura 5.2).
Deci, hemul este localizat intr-un pliu
(buzunar) hidrofob al fiecarui lant polipeptidic. Ancorarea lui la
aceste lanturi se face prin legaturi chimice coordinative cu:
histidina (in pozitia 58 si 87) de pe lanturile a si cu histidina
(in pozitia 63 si 92 de pe lantul b). Inelul imidazolic al
histidinei se leaga direct de Fe2+ al hemului, printr-o
legatura coordinativa.
Fe din molecula de Hb
se leaga hexacoordinativ de:
- cei 4 atomi de N
ai nucleului tetrapirolic prin 4 legaturi: 2 covalente (puternice) si
2 coordinative (notate 1-4)
- atomii de N ai
unui rest histidinic din lantul polipeptidic, prin 2 legaturi
coordinative (legaturile 5-6). Legatura 5 este stabila; se
realizeaza direct cu un N histidinic. Legatura 6 este labila
si va lega O2 in sangele arterial formand oxiHb, iar in sangele
venos va ramane libera .
Hb are 2 configuratii
spatiale:
oxihemoglobina - forma oxigenata sau forma relaxata. Fixarea oxigenului se
face de catre fierul din hem, in procesul de oxigenare si de
reducere, fierul ramanand bivalent. Legarea oxigenului se face la nivelul
legaturii care asigura contactul dintre fier si histidina din
pozitia 63 a lantului b (respectiv 58 pentru lantul a). In timpul oxigenarii,
pozitia spatiala a dimerilor a si b se schimba:
lanturile a se departeaza, iar lanturile b se apropie micsorand
spatiul central al tetramerului, consecinta fiind expulzarea 2,3-DPG
(produs intermediar al metabolismului glucidic eritrocitar, rezultat din
suntul Rappaport-Luebering). Aceasta forma a Hb manifesta o
mare afinitate pentru legarea O2.
deoxihemoglobina - forma
redusa, disociata (fara O2) sau tensionata,
in a carei cavitate centrala se gaseste 2,3-DPG. Trecerea
de la forma redusa la cea oxigenata nu influenteaza
valenta fierului, care ramane permanent in stare feroasa - Fe2+.
In Hb redusa, legarea fierului de nucleul imidazolic al histidinei se face
printr-o legatura de hidrogen, prin intermediul unei molecule de
apa. In OxiHb, molecula de apa este indepartata si O2
se fixeaza direct de fier.
Desi locul de legare al 2,3-DPG pe
molecula de Hb este diferit de cel al O2, intre cele 2 componente
exista o relatie reciproca de excludere. Acumularea de 2,3-DPG
in cavitatea centrala scade afinitatea Hb pentru O2, in schimb
este favorizata eliberarea O2. Expulzarea 2,3-DPG din molecula
de Hb determina cresterea afinitatii acesteia pentru O2,
fiind favorizata fixarea lui .
Rolurile hemoglobinei
Hb are trei functii, fiecare dintre ele
esentiala pentru viata: transportor de O2 de la
plamani la tesuturi, transportor (in mai mica masura)
de CO2 de la tesuturi la plamani si functia de
tamponare a sangelui pentru a preveni modificarile de pH.
Transportul oxigenului.
Cel mai important rol al Hb, transportul de O2
deriva din proprietatea Hb de a fixa reversibil oxigenul la nivelul
capilarelor pulmonare, de a-l vehicula prin sange sub protectia membranei
eritrocitare si de a-l ceda cu usurinta tesuturilor,
unde pO2 in capilarele tisulare este mica.
La o oxigenare normala, fiecare
molecula de Hb transporta 4 molecule de O2, astfel incat
oxiHb poate fi reprezentata mai corect prin simbolul Hb(O2)4
decat prin HbO2. 1g de Hb transporta 1,34-1,35 ml O2,
iar la o concentratie de Hb de 15g/dl, 100 ml de sange vor transporta 20,9
ml de O2, din care, in stare de repaus numai 4,5 ml vor fi
cedati tesuturilor. Prezenta Hb mareste de aprox. 70
de ori capacitatea sangelui de a transporta O2.
A devenit o obisnuinta sa
reprezentam afinitatea Hb pentru oxigen, dar si disocierea
(eliberarea) acestuia din oxiHb sub forma unui grafic numit curba de disociere
a oxiHb (figura 5.3) in care axa Y
reprezina procentul de Hb saturata cu O2 pentru o
solutie de Hb, la un pH de 7,4, iar axa X reprezinta presiunea partiala a oxigenului. Fixarea si disocierea O2
urmeaza o curba de aspect sigmoid, care poate fi parcursa in
ambele sensuri. Forma de S a curbei, mult mai eficienta functional
decat aspectul hiperbolic se datoreaza capacitatii diferite (in
fapt vitezei diferite) a celor 4 grupari hem de a fixa O2 pe
masura ce creste presiunea acestuia. In procesul de acceptare a
O2, gruparile hem coopereaza (interactiune hem-hem):
fixarea O2 pe o grupare hem determina modificari in
conformatia moleculei de Hb care faciliteaza oxigenarea celui de-al
doilea hem, iar acesta la randul sau creste afinitatea celui de-al
treilea hem; asa se explica de ce legarea O2 pe hemurile 3
si 4 se face mult mai rapid decat pe primul hem.
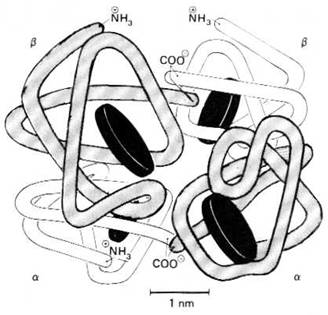
Figura
5.2. Reprezentarea unei molecule de hemoglobina A, care are 4
subunitati. Exista 2 lanturi polipeptidice α si 2
β fiecare continand un hem reprezentat printr-un disc (dupa W.F. Ganong - 1996).
Legarea
O2, dar si eliberarea lui (disocierea) depind de presiunea
partiala a gazului. La nivel pulmonar, unde pO2 este de 100 mmHg, Hb este
aproape complet saturata si transformata in oxiHb. La nivel
tisular, unde pO2 este de 40mmHg o buna parte din oxiHb se
disociaza, eliberand O2 (procentul de saturare este de 70%),
deci scaderea pO2 la nivelul tesuturilor favorizeaza
disocierea.
Cand pO2 este sub 20 mmHg, Hb are o
foarte slaba afinitate pentru oxigen, iar partea initiala a
curbei (presiuni mici) este usor inclinata.
Pentru acceptarea primelor cantitati
de O2 este necesara o pO2 peste 20 mmHg, valoare de
la care afinitatea Hb pentru oxigen creste substantial. Intre 20-30
mmHg, curba devine aproape verticala (la variatii mici ale pO2
exista variatii mari ale saturatiei Hb cu O2),
indicand ca Hb poate ceda rapid O2 fixat. La pO2
peste 70 mmHg, acceptarea se va face mai greu, curba devine mai lina, in
platou. La o pO2 de 100 mmHg (capilarele alveolare), saturarea cu
oxigen a Hb este de 97,5%.
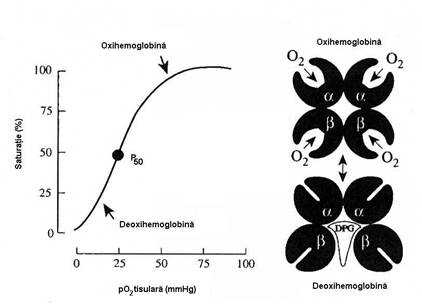
Figura nr. 5.3 Curba de fixare-disociere a Hb (stanga) si
modificarile spatiale ale moleculei de Hb in timpul fixarii
si cedarii O2
In afara interactiunii hem-hem si a
dependentei de pO2, capacitatea Hb de a fixa si ceda O2
mai este influentata si de alti factori intra sau
extraeritrocitari (tabel 5.2 si figura 5.4):
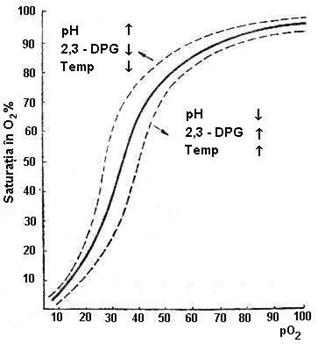
- cresterea concentratiei de CO2. La nivelul
capilarelor tisulare CO2 trece din plasma in eritrocit, unde
legand o molecula de apa (in prezenta anhidrazei carbonice) se
transforma in acid carbonic (H2CO3), care
disociaza rapid in H+ si HCO3-.
Acumularea de protoni scade pH-ul intracelular, care scade afinitatea Hb pentru
O2 si faciliteaza disocierea acestuia (deplaseaza la
dreapta curba de disociere).
- 2,3 - DPG modifica curba de fixare-disociere: prin expulzarea
sa din cavitatea centrala a deoxiHb favorizeaza legarea O2,
iar prin legarea de Hb favorizeaza cedarea O2 catre
tesuturi (deci cresterea cantitatii de 2,3-DPG
deplaseaza curba la dreapta). Actioneaza prin modificarea
pH-ului intracelular (scade pH-ul eritrocitului fata de cel
plasmatic).
- valoarea pH-ului sanguin (efectul Bohr): acumularea de
metaboliti acizi la nivel tisular scade pH-ul, favorizand disocierea oxiHb
(curba la drepta). La nivelul capilarelor pulmonare, unde pH-ul este mai
crescut creste afinitatea pentru O2, favorizandu-se fixarea.
- temperatura sangelui: solubilitatea unui gaz intr-un lichid este
invers proportionala cu temperatura lichidului la presiune
constanta. Cand temperatura sangelui creste, sensul reactiei: Hb
+O2 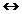 HbO2 se
deplaseaza spre stanga, deoarece scade concentratia oxigenului.
Aceasta echivaleaza cu cresterea disocierii (si scaderea
afinitatii), adica cu deplasarea la dreapta a curbei.
HbO2 se
deplaseaza spre stanga, deoarece scade concentratia oxigenului.
Aceasta echivaleaza cu cresterea disocierii (si scaderea
afinitatii), adica cu deplasarea la dreapta a curbei.
|
Parametri
|
Deplasarea
curbei este determinata de:
|
|
Cresterea
|
Scaderea
|
|
Temperatura
sangelui
|
D
|
S
|
|
pH
|
S
|
D
|
|
2,3
- DPG eritrocitar
|
D
|
S
|
|
CO2
|
D
|
S
|
Tabelul nr.5.2. Factorii care determina
deplasarea curbei la dreapta (D), asociati cu scaderea
afinitatii pentru fixarea O2 (disociere) si la stanga
(S), asociati cu cresterea afinitatii pentru fixarea O2.
Figura
5.4. Deplasarea curbei de disociere a Hb sub actiunea unor factori
intra si extraeritrocitari
|
|
Transportul bioxidului de carbon
In afara de
transportul O2, Hb participa si la transportul CO2
de la tesuturi la plamani, de unde este eliminat prin expiratie.
Aproximativ 70% din CO2 produs la nivel tisular este transportat
prin plasma, in cea mai mare parte sub forma de bicarbonat, iar 30%
este vehiculat de eritrocit: 25% sub forma de bicarbonat, 2% sub
forma neionizata (ambele forme dizolvate in apa eritrocitara)
si restul de 3% sub forma de carbaminHb. Interventia
eritrocitului in transportul CO2 are drept consecinta nu
numai eliminarea gazului, ci si implicarea in mentinerea
balantei acido-bazice, prin rolul de sistem tampon jucat atat de oxiHb,
cit si de deoxiHb.
Rolul Hb in balanta acido-bazica
Eritrocitul poseda
trei sisteme tampon, dintre care doua sunt realizate prin intermediul
hemoglobinei: HHb/KHb (Hb redusa/hemoglobinat de K) si HbO2/
KHbO2 (oxi Hb/oxihemoglobinat de K). Hb, ca orice proteina este
o substanta amfotera, care poate actiona ca acid
(doneaza protoni) prin gruparile carboxil, sau ca baza
(accepta protoni) prin gruparile amino. Hemoglobina neoxigenata
(Hb-) este o baza mai puternica decat oxiHb, ea putand
sa accepte si sa neutralizeze mai usor protonii
rezultati ca urmare a cresterii nivelului sanguin al CO2.
Tipuri de hemoglobine
normale (fizIologice)
Hb A1
(a2b2) Reprezinta componenta
majora din eritrocitele adultului normal. Are in structura sa doua
lanturi a si doua b, care difera prin structura lor primara.
Principale caracteristici sunt: mobilitate mare electroforetica, punct
izoelectric (pH 6,87) si afinitate crescuta pentru O2.
Cele mai multe proprietati fizice si chimice referitoare la Hb
au fost obtinute prin studiul Hb A1.
HbF a2g2) reprezinta componenta majora a Hb in perioada
dezvoltarii fetale. In sangele din cordonul ombilical si la nou
nascut este in proportie de 70-80% din totalul Hb. Principala
caracteristica o reprezinta rezistenta crescuta la
denaturarea alcalina, pe aceasta proprietate bazandu-se si
determinarea ei cantitativa. O alta caracteristica a HbF este
afinitatea crescuta pentru O2 in raport cu Hb A1,
acest fapt datorandu-se faptului ca HbF leaga 2,3 DPG mai greu decat Hb A1.
Datorita acestei caracteristici O2 de pe Hb A1 din
eritrocitele materne poate trece pe Hb F din eritrocitele fetale (la nivelul
capilarelor vilozitatilor coriale ale placentei). Valori mari de HbF
se intalnesc fiziologic in sarcina si patologic in anemii severe: b talasemia hetero
si homozigota, anemia Biermer grava sau in leucemii.
In afara acestor Hb fiziologice, care au
structuri proprii, determinate genetic se mai cunosc si Hb minore,
nedeterminate genetic, care sunt de fapt Hb normale modificate in vivo sau in vitro. Dintre acestea, cea mai importanta este Hb A1c
care are atasat de lantul a sau d un rest glucidic. Acest tip de Hb creste in
eritrocitele senescente si in diabetul zaharat, direct proportional
cu nivelul mediu al glicemiei.
|
Tipul de hemoglobina
|
Structura moleculara
|
Varsta la care apare
|
Proportia (%)
|
|
Nou nascuti
|
Adulti
|
|
Portland
|
z2g2
|
Numai in viata embrionara
|
|
|
|
Grower I
|
z2e2
|
|
|
|
Grower II
|
a2e2
|
|
|
|
Fetala (F)
|
a2g2
|
Si la nou nascut si la adult
|
|
< 1
|
|
A1
|
a2b2
|
|
|
|
A2
|
a2d2
|
< 0,5
|
|
Tabelul
5.3. Tipuri de hemoglobine fiziologice.
Derivatii de Hb In
afara O2, Hb se poate combina si cu alte substante pentru
care manifesta o afinitate crescuta, formand compusi instabili
sau stabili, lipsiti de capacitatea de a fixa si transporta O2.
- Oxihemoglobina (vezi mai sus)
▪ Carbaminhemoglobina sau carbohemoglobina este compusul
fiziologic in care Hb leaga CO2. Legarea acestuia nu se face la
nivelul fierului, ca in cazul O2, ci la gruparile aminice
libere ale globinei, rezultand un compus carboaminic, forma sub care se
transporta o parte din CO2 de la tesuturi la plamani,
pentru a fi eliminat. Restul de CO2 este eliminat fie sub forma
dizolvata in plasma (sub 5%), fie legat de bicarbonatul plasmatic sau
de proteine. Cu cat Hb este mai desaturata (a eliberat O2), cu
atat legarea CO2 este mai facila si invers, eliberarea CO2
este favorizata de reoxigenarea Hb. In carbaminhemoglobina, fierul
hemului nu-si modifica valenta (ramane bivalent).
Derivatii patologici
de Hb
▪ Carboxihemoglobina (COHb). Este derivatul de Hb format
prin legarea monoxidului de carbon. Este un compus reversibil, in care CO se
leaga direct de Fe2+, in locul O2. (valenta
fierului nu se modifica). Hb are o afinitate de 200 de ori mai mare pentru
CO decat pentru O2, ceea ce explica rapiditatea instalarii
si gravitatea intoxicatiei cu CO, chiar la concentratii mici ale
CO in aerul inspirat. In plus, viteza de disociere a COHb este lenta,
cantitatea scazand la jumatate in 4 ore.
In mod normal in sange se gaseste o
cantitate sub 2% de COHb, care poate creste pana la 4% la
fumatori si persoanele care traiesc in zone intens poluate.
Sursele de intoxicatie sunt reprezentate
de arderile incomplete ale hidrocarburilor: gazele de esapament, sobe cu
reglaj sau tiraj deficitar, furnale etc. Intoxicatia este posibila
deoarece CO este un gaz fara miros sau culoare.
Primele semne de intoxicatie apar chiar
la concentratii de 10-20% si constau in cefalee, ameteli,
iritabilitate, greata, dispnee, culoare
rosie aprinsa a tegumentelor
si mucoaselor, semnul cel mai caracteristic al
intoxicatiei cu monoxid de carbon. La concentratii de 50%,
intoxicatia este grava, pacientul prezentand voma, convulsii,
coma si se poate instala moartea. Tratamentul consta in
scoaterea pacientului din mediul toxic si administrarea de O2
hiperbar (2-3 atm.), care creste viteza de disociere a COHb, timpul de
injumatatire scazand de la 4 h, la 1h. Oxigenoterapia
hiperbara creste pO2 si implicit cantitatea de O2
solvit in plasma, care va suplini partial absenta Hb,
facand posibila salvarea pacientului; adesea, din pacate, in
intoxicatiile severe raman sechele ale SNC sau cordului.
▪
Methemoglobina rezulta in urma oxidarii Fe2+ (feros)
al Hb si trecerea lui in Fe3+ (feric). Poate lega O2,
dar printr-o legatura stabila - nu-l mai poate ceda, ceea ce
face ca MetHb sa nu poata indeplini rolul de transportor de O2.
Cantitati mici, sub 5% de MetHb apar in sange si in
conditii fiziologice, ca rezultat al oxidarii Hb sub actiunea O2
transportat, dar sunt rapid reduse de catre sistemele reducatoare ale
eritrocitului, intre care NADH-methemoglobin reductaza are rolul cel mai
important. Cresterea concentratiei la 10-25% determina
aparitia cianozei, dar nu necesita tratament. La o concentratie
de 35% apar: cefalee, dispnee, oboseala marcata, iar la 70% se poate
produce moartea. Sangele care contine cantitati mari de metHb
are o culoare bruna, iar examinarea spectroscopica indica o
banda de absorbtie caracteristica: in rosu, la 620-630 nm,
cate o banda ingusta in galben si in verde si o banda
lata in albastru.
MetHb poate aparea in
urmatoarele circumstante:
-
expunerea eritrocitelor la agenti oxidanti-nitriti,
coloranti anilinici (folositi si in industria alimentara),
naftalen etc. Intoxicatia cu nitritii proveniti din apa
poate fi mortala la sugari, daca de la primele semne de cianoza
nu se intervine terapeutic prin administrare de substante reducatoare:
albastru de metilen (este un foarte bun donor de electroni), administrat oral
sau intravenos in intoxicatiile grave, la care se adauga vitamina C,
de regula i.v. La toate intoxicatiile cu MetHb, indiferent de cauza
acestora, antidotul este albastrul de metilen.
-
prezenta hemoglobinei anormale M, la care substitutia unor aminoacizi
in lantul globinic determina oxidarea Fe2;
-
in caz de deficienta congenitala de NADH-methemoglobin
reductaza, care, in mod normal reduce micile cantitati de metHb
care apar fiziologic in eritrocite. Indivizii cu o astfel de patologie sunt
tratati cronic cu albastru de metilen, care va colora pielea si
urina; in lumea medicala sunt cunoscuti ca "oamenii albastri"
sau "familii albastre", deoarece deficienta de NADH-methemoglobin reductaza
este congenitala (Trost, C, 1982).
▪ Sulfhemoglobina
(sulfHb) este un derivat hemoglobinic, care spre deosebire de COHb si
MetHb nu poate fi convertita in Hb. Odata formata nu poate fi
indepartata din organism decat prin flebotomie. Apare in urma
utilizarii cronice a unor droguri (medicamente) ca acetanilida sau
fenacetina, la unele persoane cu constipatie cronica si
producere in exces de hidrogen sulfurat (cianoza enterogena) si
foarte rar este o maladie ereditara. Sangele care contine SulfHb are
o culoare bruna-mov.
Anomalii
ale sintezei globinei. Hemoglobinopatiile
Hemoglobinopatiile
sunt boli ereditare in care este afectata rata de sinteza sau
structura globinei, exprimate clinic si biologic, cel mai frecvent sub
forma de anemie hemolitica, rareori sub forma de cianoza
sau poliglobulie. Exista si anomalii dobandite ale Hb, de obicei sub
actiunea unor substante toxice, care nu sunt insa acceptate de
multi autori, ca facand parte din aceste entitati
nosologice.
Se
disting doua mari grupe de hemoglobinopatii : cantitative caracterizate
prin blocarea partiala sau totala a ratei de sinteza a
unuia sau mai multor lanturi polipeptidice ale globinei si calitative
in care anomalia sau mutatia genetica duce la sinteza unor Hb
anormale din punct de vedere chimic. Gravitatea hemoglobinopatiilor este
variabila, de la anomalii de laborator, asimptomatice pana la
afectiuni cu sfarsit letal in uter sau in prima copilarie.
Hemoglobinopatii cantitative - Sindroamele talasemice (anemia
mediteraneana)
▪ Talasemia b In acest tip de hemoglobinopatie, anomalia
genetica produce blocarea partiala sau totala a sintezei
lanturilor b. Are o forma majora
caracterizata prin absenta completa a lanturilor b (deficit homozigot) numita si anemia Cooley. Deoarece Hb A1
nu se poate sintetiza, ea este inlocuita cu HbA2 si HbF.
Din punct de vedere clinic se exprima prin anemie hemolitica
severa, dependenta de transfuzii, hepato-splenomegalie,
modificari osoase, deces in prima sau a doua decada a vietii.
Frotiul arata o anemie, hipocroma, anizopoikilocitoza, cu hematii
in tinta, in picatura, ovalocitoza, schizocite (Kobbi
S,1998).
Talasemiile b ocupa primul loc
in cadrul hemoglobinopatiilor in intreaga lume, atat prin incidenta
crescuta, cat si prin gravitate. In Europa, incidenta
maxima este in tarile din bazinul Mediteranean (Cipru, Italia,
Grecia).
▪ Talasemia a se caracterizeaza prin absenta sau scaderea sintezei
lanturilor a
si cresterea sintezei
de non a.
Lanturile a sunt inlocuite cu b b) in Hb H
(raspandita in Africa, Asia de S-E, bazinul mediteranean) sau cu g g) in Hb Barth.
Hemoglobinopatii calitative
Sunt
datorate unor mutatii genetice, care se exprima prin: inlocuirea
(substitutia) unor aminoacizi; prin pierderea (deletia) unuia sau mai
multor aminoacizi sau prin crossing-over.
▪ Prin substitutie:
- siclemia sau hemoglobinopatia S (drepanocitoza, anemia
falciforma, anemia cu hematii in secera). Se datoreaza
inlocuirii acidului glutamic din pozitia 6 de pe lantul b cu valina, care modifica configuratia
spatiala a lanturilor, scade solubilitatea Hb, determina
formarea de cristale de Hb (tactoizi), care la randul lor induc modificarea de
forma a eritrocitelor (drepanocit, secera). Se transmite autozomal (nelegat de sex).
Boala se caracterizeaza prin anemie hemolitica cronica si
fenomene de ocluzie vasculara determinate de transformarea
drepanocitara a eritrocitelor. Incidenta acestei boli este
maxima in zona ecuatoriala a Africii, in Peninsula Arabica,
sudul Indiei, Madagascar si populatia de culoare din cele doua
Americi. Forma heterozigota (AS) confera rezistenta la
malarie (Kobbi, S, 1998).
- alte hemoglobinopatii in care substitutia unui aminoacid
intereseaza tot lantul b sunt:
hemoglobinopatia C (b6 Glu Lys), in care
prezenta acestui tip de Hb determina rigidizarea eritrocitelor si
sechestrarea lor splenica; hemoglobinopatia D, cu cel putin 6
variante structurale, toate prezente pe lantul b; hemoglobinopatia E (b26 Glu Lys). In
hemoglobinopatia O (HbX) substitutia intereseaza atat lanturile a cat si b.
Lys), in care
prezenta acestui tip de Hb determina rigidizarea eritrocitelor si
sechestrarea lor splenica; hemoglobinopatia D, cu cel putin 6
variante structurale, toate prezente pe lantul b; hemoglobinopatia E (b26 Glu Lys). In
hemoglobinopatia O (HbX) substitutia intereseaza atat lanturile a cat si b.
▪ Prin deletie - hemoglobine instabile. Intereseaza
unul sau ambele lanturi polipeptidice si se caracterizeaza prin
anemie hemolitica si prezenta corpilor Heinz in eritrocite: Hb
Zürich, Hb Bucuresti, Hb Tacoma etc..

