RECOMBINAREA GENETICA
LA EUCARIOTE
Recombinarea genetica
este procesul prin care are loc transferul intra- sau intermolecular a unor
secvente de ADN, avand ca rezultat modificari in
inlantuirea genelor sau a unor parti din gene. Se poate produce astfel, reasortarea unor
nucleotide la nivelul aceleiasi molecule de ADN sau intre molecule
separate, rezultand, in final, una sau doua molecule recombinate.
La
eucariote procesul de recombinare genetica este legat de fenomenul
sexualitatii si se realizeaza in special in cursul meiozei,
fiind conditionata de realizarea contactelor intre cromosomi
urmata de disjunctia independenta a perechilor de cromosomi
(bivalenti). In cazul organismelor eucariote au fost descrise 3 tipuri de
recombinare genetica:
recombinarea intracromosomala - prin
crossing-over (schimbul reciproc de segmente cromosomale);
recombinarea intercromosomala - prin
disjunctia independenta a cromosomilor omologi;
recombinarea genetica
nereciproca - prin conversie genetica.
Recombinarea intracromosomala (crossing-over) se realizeazaatat in meioza cat
si in mitoza.
Ea poate avea loc intergenic, cat si intragenic.
Recombinarea intragenica este un fenomen rar. Acest fenomen manifesta interferenta
negativa care determina cresterea probabilitatii ca un
al 2-lea crossing-over sa aiba loc in apropierea primului.
Formarea bivalentilor in
meioza presupune participarea a 2 cromosomi de origine diferita
(materna si paterna), intre care se stabilesc contacte
intercromatidice (sinapse) ce conduce la formarea unei structuri specializate
numita complex sinaptinemal.
Importanta complexului
sinaptinemal in desfasurarea recombinarii genetice
intracromosomale este dovedita de faptul ca la organismele la care nu
se produce acest complex, nu are loc procesul de crossing-over.
Formarea chiasmelor este
dependenta de sinteze proteice ce au loc la sfarsitul zigotenului.
Procesul de recombinare genetica implica ruperea si reunirea
cromatidelor participante, fapt ce conduce la un schimb reciproc de segmente de
ADN de dimensiuni egale si la aparitia de cromosomi (iar apoi de
gameti) recombinati (Fig. 1).
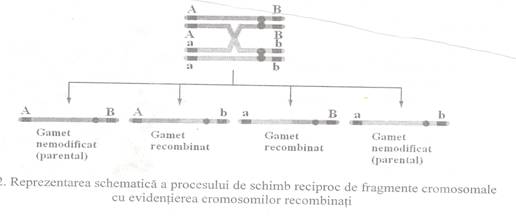
Fenomenul de crossing-over se realizeaza mai mult sau mai
putin randomic, de-a lungul perechilor de cromosomi omologi. Astfel, probabilitatea realizarii recombinarii
intre 2 loci creste odata cu distanta dintre acestia pe
cromosomi.
Deoarece
procesul de crossing-over afecteaza numai 2 dintre cromatidele
bivalentilor, doar 50 % dintre produsii meiozei sunt de tip
recombinat, restul de 50 % sunt de tip parental.
S-a
constatat ca intre genele plasate pe cromosomii omologi, in afara de
crossing-over simplu, pot aparea si crossing-overe multiple (Fig.2).
Daca procesul de recombinare are loc la nivelul unor gene heterozigotze,
indiferent de numarul de crossing-overe ce se realizeaza, procentul
de recombinare nu depaseste 50 %.
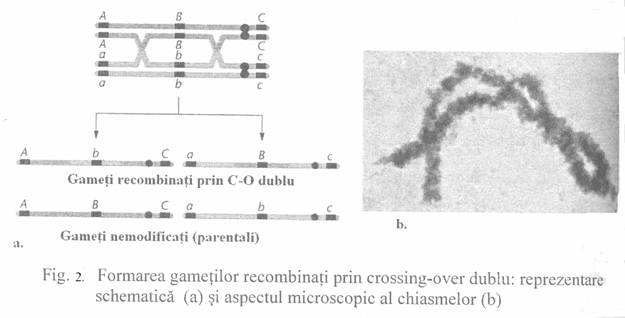
Procesul
recombinarii genetice poate avea loc si in celulele somatice.
Schimburile intercromatidice (sister chromatid
exchange) au fost evidentiate mai ales in celulele mamaliene aflate in
cultura in vitro, ele nefiind
urmate, de obicei, de modificari fenotipice deoarece cromatidele implicate
sunt identice.
Fenomenul recombinarii mitotice a
fost studiat in special in cazul fungilor (Aspergillus
nidulans, S. cerevisiae) la care in ciclul de viata
predomina haplofaza.
Recombinarea
mitotica poate avea loc in cursul metafazei si presupune asocierea
cromosomilor omologi si realizarea schimbului genetic inainte de
diviziunea centromerului.
Procesul de
crossing-over se poate produce si la nivelul aceleiasi gene
(crossing-over intragenic).
In
mecanismul molecular al recombinarii genetice de tip crossing-over, sunt
implicate proteine specifice, procesul recombinarii fiind asociat cu cel
de reparare genetica.
Recombinarea intercromosomala are loc prin disjunctia
independenta a perechilor de cromosomi care reprezinta suportul
material pentru disjunctia perechilor de factori ereditari.
Segregarea
independenta are loc la trecerea celulelor sexuale de la metafaza I la
anafaza I, cand are loc asezarea intamplatoare a cromosomilor de
origine materna sau paterna deasupra sau dedesubtul planului
ecuatorial al placii metafazice.
Acest
comportament al cromosomilor de origine matena, care se imperecheaza
cu cei de origine paterna si apoi separarea perechilor respective, a
fost comparat de catre Muller cu formarea perechilor de dansatori in
timpul dansului si separarea lor la sfarsitul acestuia. Din
aceasta cauza, Muller a denumit acest comportament dansul
cromosomilor . Cu cat numarul cromosomilor si a genelor
continute este mai mare cu atat numarul de combinatii posibile
de gameti si de organisme este mai mare. Posibilitatea ca un organism
sa fie asemanator cu altul este de (1/2)2n, in care n
=numar de perechi de cromosomi, iar 2n reprezinta numarul total
de cromosomi din complementul speciei respective.
Conversia genica este o recombinare nereciproca. In
cadrul conversiei genice, un segment din molecula de ADN a unei cromatide se desprinde
si se ataseaza de alta cromatida. Este posibil, in acest mod, ca un
cromozom sa contina o gena in doua exemplare (daca transferul s-a realizat
intre cromozomi omologi), iar altul o gena in minus. Are loc cu o frecventa mai
mare in cursul diviziunii meiotice dar poate avea loc si in celulele somatice. Ea a fost evidentiata prin analiza
tetradelor la ciupercile din genurile Saccharomyces,
Aspergillus,
Neurospora unde apar abateri de la raportul normal de segregare
alelica, ca si cum se produce o transformare a alelei A in a (Fig. 3).
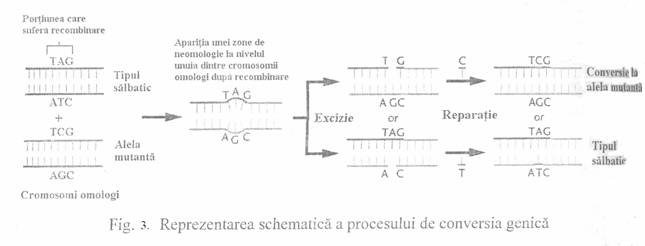
Frecventa
recombinarii genetice nereciproce poate creste de la un capat la
altul al unui locus genic, fenomen numit polarizarea
conversiei. Astfel , la fungi, in locul raportului de segregare normal de
4 : 4, se intalnesv valori
diferite ale acestui raport: 6 : 2, 5 : 3, 7: 1 sau chiar 8 :0. fenomenul
conversiei genice a fost evidentiat si la plante, in cazul unor gene
ce intervin in determinismul pigmentatiei, precum si la virusuri (in
special bacteriofagi). Unii autori considera ca, de fapt, conversia
genica este consecinta unui proces de recombinare, in urma
caruia apar nepotriviri ale unor baze azotate situate pe cromatide
omoloage. Aceste zone de nepotrivire sunt reparate prin excizie si prin
adaugarea nucleotidei corespunzatoare. Prin reparatie poate
reaparea astfel, tipul normal.
Curs 12-
Genetica
RECOMBINAREA GENETICA
LA PROCARIOTE
La
procariote au fost descrise mai multe cai de transfer de material genetic: transformare genetica, conjugare,
sexductie si transductie, asociate cu mecanisme ce
asigura integrarea noii informatii genetice in genomul gazdei.
Indiferent de natura sa, transferul de gene reprezinta un fenomen in care
moeculele informationale isi schimba gazda, avand de traversat
doua bariere celulare: una la iesire din celula donatoare si
alta la intrarea in celula receptoare.
- Transformarea genetica
Fenomenul transformarii
genetice a fost descoperit in 1928 de catre Griffith, in urma
experimentelor realizate cu pneumococi asupra soarecilor. Ulterior ,
Avery, Mac Leod si McCarthy, au descoperit, in 1944, ca agentul transformant
este reprezentat de ADN izolat din celulele bacteriene.
Transformarea genetica la bacterii presupune transferul unui fragment
de ADN (cromosomal sau plasmidial) de la o bacterie donatoare si
incorporarea acestuia in genomul unei bacterii receptoare.
Asemenea fragmente de ADN
sunt, in general, de dimensiuni mari (in medie de 20.000 perechi de
nucleotide), putand contine gene utile care, patrunse in bacteria
receptoare, pot inlocui printr-un proces de recombinare o secventa de
nucleotide omoloaga din genomul acesteia. Informatia genetica
nou integrata in cromosomul bacteriei receptoare este transmisa
stabil de-a lungul generatiilor.
Fenomenul de transformare
genetica a fost evidentiat prima data la pneumococul Diploccocus pneumoniae si ulterior
la Bacillus subtilis, Escherichia coli,
Haemophilus influenzae, Staphylococcus sp., etc.
Pentru a putea
prelua ADN exogen din mediul extracelular, celulele receptoare trebuie sa
manifeste o
anumita stare specifica numita stare de competenta. Aceasta presupune
ca peretele celular si membrana celulara ale celulei bacteriene
receptoare sa fie permeabile pentru ADN exogen, iar celula sa se afle
intr-o anumita faza a ciclului de viata care sa
permita acceptarea acestuia. In cursul realizarii competentei
are loc activarea unor receptori specifici localizati la nivelul peretelui
celular cu rol de legare a ADN exoged la suprafata celulara.
Numarul receptorilor activi, variaza de la o specie bacteriana
la alta, de exemplu: la Streptococcus
au fost identificati 80 de receptori, la Bacillus subtilis - 50, iar la Haemophilus
doar 4 receptori.
La
unele specii bacteriene, starea de competenta este o stare
naturala, fiziologica, aparand intr-o anumita etapa a
ciclului de viata si dureaza un anumit interval de timp. De
exemplu, la Azotobacter, Staphylococcus,
Pneumococcus competenta este maxima pe parcursul fazei
logaritmice, in timp ce la alte specii - Bacillus,
Pseudomonas, Methylobacterium- competenta este maxima la trecerea
de la faza logaritmica la cea stationara.
In
cazul altor specii, starea de competenta poate fi indusa prin
tratamente specifice (soc de temperatura, adaugarea unor ioni
etc.).
Intr-o
populatie bacteriana, proportia celulelor capabile de a prelua
ADN exogen poate fi estimata pe baza frecventei de transformare,
urmarind un anumit marker genetic. Pentru a se obtine o
frecventa ridicata de integrare/ transformare, este necesar ca
ADN transformant sa provina de la o tulpina bacteriana
inrudita (ADN omolog). In cazul utilizarii unor organisme neinrudite
(ADN heterolog), eficienta procesului de transformare este foarte
scazuta.
Multe
bacterii se pot liza in mod natural, mai ales in timpul etapelor finale ale
ciclului celular. In aceste conditii, ADN este eliberat in mediu
extracelular de unde poate fi preluat de alte bacterii aflate in aceeasi
populatie.
La bacterii, procesul natural de
transformare genetica (cu fragmente de ADN cromosomal eliberat prin metoda
lizei celulelor bacteriene) se realizeaza in mai multe etape:
- legarea reversibila (adsorbtia) a moleculelor de ADN dublu catenar,
la nivelul situsurilor receptoare aflate pe suprafata celulara;
- preluarea ADN exogen de catre celulele competente;
- convertirea ADN dublu catenar la forma
monocatenara,
prin degradarea uneia dintre catenele moleculei initiale;
- integrarea segmentului de ADN monocatenar in cromosomul celulei receptoare;
- segregarea si exprimarea fenotipica
a noii
informatii genetice integrate in genomul celulei gazda.
Procesul de transformare este
destul de raspandit in natura, mai ales in cazul tulpinilor
bacteriene inrudite, el jucand un rol important in evolutia bacteriilor.
Aceasta concluzie este intarita si de observatia
ca transferul de gene prin transformare genetica are loc nu numai pe
orizontala (intre bacterii inrudite) ci si intre grupe de organisme
neinrudite (fenomen ce explica, de exemplu, raspndirea
rezistentei la antibiotice).
- Conjugarea bacteriana
Conjugarea bacteriana
reprezinta un mod de transfer unidirectional de ADN de la o
celula donatoare la o celula
receptoare prin intermediul unei legaturi intercelulare directe (pil de
sex) si conditonat de prezenta in bacteria donatoare a unui
element genetoic specializat numit conjugon.
Fenomenul
conjugarii a fost descoperit in 1946 (de catre Lederberg si
Tatum) in urma experimentelor cu tulpini de Escherichia
coli ce contineau sau nu o plasmida specifica notata F.
Din acest punct de vedere, celulele bacteriene care contin plasmida F (
factorul F) sunt notate F +, iar cele care nu contin
aceasta plasmida sunt notate F -. In cazul bacteriilor
Gram negative, in afara de plasmida F, functiile necesare procesului
de conjugare se gasesc la nivelul mai multor plasmide. De exemplu, in
cazul plasmidelor de rezistenta la antibiotice (plasmida RP4) si
plasmidele Col ce contin
determinanti genetici pentru sinteza colicinelor.
Factorul F
este alcatuit din ADN dublu catenar circular cu o lungime de 94,5 kb
si in numar de 1-2 copii/celula. Transferul prin conjugare al
acestor plasmide se datoreaza prezentei unei regiuni specifice,
regiunea tra, ce reprezinta cca
30 % din lungimea plasmidei F. Cu exceptia fectorului F, celelalte
plasmide conjugative pot include si alte tipuri de gene: pentru
rezistenta la antibiotice, toleranta la metale grele,
virulenta patogenitate etc.
Studiile asupra procesului de conjugare
efectuate la E. coli , au permis
stabilirea etapelor acestuia:
Formarea si stabilirea agregatelor de conjugare;
Pregatirea ADN pentru transfer
Transferul ADN conjugativ
Refacerea structurii circulare si dublu catenare a ADN transferat.
In prima etapa
are loc sinteza de catre celula donatoare a pililor de sex - structuri
specializate, tubulare, localizate la suprafata celulara,
alcatuite din molecule de pilina (proteina a carei
sinteza este codificata de gene plasmidiale).
A 2-a etapa a
conjugarii presupune interventia unei proteine, codificata de
genele plasmidiale, care are rolul de a
realiza o crestatura monocatenara la nivelul genei oriT (originea transferului). La nivelul capatului 5' al catenei de ADN crestata, se leaga
o proteina specifica, codificata tot de gene plasmidiale de la
nivelul regiunii tra. Are loc apoi, transferul catenei crestate,
incepand cu capatul 5' , transferul fiind cuplat cu procesul de replicare
al plasmidei, conform modelului cercului rotativ. In acest fel, pe
masura ce procesul de replicare avanseaza, una dintre catenele
vechi (cea crestata) se deplaseaza prin pilul de sex in celula
receptoare (Fig. 1).
In acelasi timp, in
celula donatoare are loc restabilirea structurii factorului F. la nivel
populatiei de bacterii, consecinta transferului factorului F este masculinizarea acesteia, adica se
produce generalizarea caracterului F +al celulelor bacteriene.
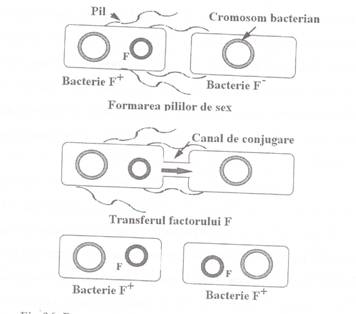
Fig. 1. Reprezentarea schematica a
procesului de conjugare
Un tip special de conjugare
este cel realizat intre celulele bacteriene care contin factorul F
integrat in cromosom (celule de tip Hfr) si celulele de tip F - (Fig. 2). Integrarea
factorului F in cromosomul bacterian nu se realizeaza la intamplare , ci
la nivelul unor situsuri ce prezinta omologie intre cele doua tipuri
de molecule. Celulele rezultate in urma unui asemenea proces de integrare , au
primit denumirea de celule de tip Hfr (High
frequency of recombination).
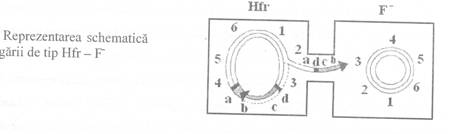
Modalitatea specifica de
integrare a plasmidei F in cromosomul bacterian, face ca secventa oriT sa fie localizata aproape de una dintre marginile plasmidei
linearizate si integrate la capatul acesteia, consecinta fiind
transferul specific al ADN monocatenar in celula receptoare.
In a 3-a etapa,
in cazul transferului dintre bacteriile F + si F -, dupa crestarea
monocatenara la nivelul secventei OriT,
incepe transferul AND monocatenar reprezentat de o scurta
secventa din plasmida F si apoi de o copie a AND cromosomal al
gazdei (Fig. 2). Cea mai mare parte a plasmidei F ramane la nivelul celuilalt
capat al cromosomului, si doar foarte rar este transferata in
celula acceptoare. Cantitatea de informatie genetica
transferata, proprie celulei donatoare, este direct
proportionala cu timpul in care cele doua celule au stat in
contact (cu cat contactul este mai lung, cu atat segmentul de ADN transferat va
fi mai mare). Astfel, pentru transferul complet al ADN cromosomal la E. coli sunt necesare aproximativ 90-120
minute de contact intre celule, in timp ce transferul plasmidei F din bacteria F +
intr-o bacterie F - dureaza 2-3 min.
Procesul de transfer de
segmente cromosomale din bacteria donor Hfr in cea acceptoare F -
este urmat de cele mai multe ori de transformare genetica (fenomen
controlat de o serie de enzime sintetizate de gazda) ceea ce inseamna
ca portiuni din respectivul segment se integreaza in genomul
noii gazde fiind mentinut si transmis constant in generatiile
urmatoare.
Procesul de conjugare dintre
celulele Hfr si celulele de tip F - serveste la
realizarea hartilor cromosomale la bacterii , distanta dintre
gene (dispuse in ordine pe segmentul transferat) exprimandu-se in
unitati de timp (minute).
Transferul de gene cromosomale
bacteriene mediat de plasmidele de sex este numit sexductie. In urma conjugarii F + x F - se
obtin doar celule de tip F + , care sunt partial ' diploide ', continand alaturi
de genele proprii si gene similare provenite de la bacteria donatoare.
3. Transductia fagica
Transductia
fagica este procesul prin care un fragment din cromosomul bacterian este
transferat de la o bacterie la alta prin intermediul capsidei (invelisului
proteic) anumitor bacteriofagi temperati. Fenomenul a fost descris
initial la bacteriofagi specifici unor tulpini de Salmonella typhimurium, iar ulterior el a fost mai bine studiat in
cazul fagului λ specific tulpinilor
de E. Coli . In functie de
structura genetica a fagilor transductori si de mecanismul de formare
a materialului genetic al particulri fagice, au fost descrise doua tipuri
de transductie fagica: transductie specializata si transductie
generalizata.
Bacteiofagii
ce realizeaza transductia generalizata produc particule ce
contin, in cea mai mare parte, ADN provenit din bacteria gazda (din
orice parte a cromosomului) si doar o foarte mica parte genom fagic.
In cazul fagilor ce determina
transductia specializata, ei produc particule ce contin atat
gene fagice (majoritare) cat si gene cromosomale bacteriene provenite
dintr-o anumita regiune a cromosomului bacterian. Cel mai cunoscut
bacteriofag ce realizeaza transductie specializata este fagul
λ. In forma lui naturala, fagul λ poate psrelua, ca fag
transductor, doar scurte secvente de ADN din genomul gazdei in care s-a
multiplicat (dupa ce a parcurs ciclul lizogen), dimensiunea acestora fiind
limitata de capsida fagica. Secventele pe care le poate prelua
fagul λ sunt foarte precise.
Tranductia generalizata este
mediata de fagi virulenti precum si de anumiti fagi
temperati ai caror genom nu se integreaza la nivelul unor
situsuri specifice din cromosomul gazdei (de exemplu fagul P1).

