Introducere
Lucrarea "
Morfologia microscopica normala la animalelor domestice"(cu elemente
de embriologie) este destinata in
principal studentilor in medicina veterinara, dar este
utila si altor specialisti care lucreaza pe animalele
domestice, precum medici veterinari,
ingineri zootehnisti, biotehnologi si biologi.
Preluand datele de microscopie optica,
microscopie electronica, embriologie si imbinandu-le cu rezultatele
stiintifice obtinute in activitatea, de peste treizeci de ani
din domeniul morfologiei normale a autorului, s-a elaborat o lucrare
unitara, logic structurata si ancorata in
realitatile clinicii si practicii veterinare. Lucrarea prezinta nivelul molecular, celular,
tisular si sistemic de organizare a organismului animal, impreuna cu
notiunile de embriologie aferente.
Se
urmareste, astfel, ca instruirea actualilor studenti, viitori
medici vetrinari sa se realizeze metodic, de la simplu la complex, incat
insusirea cunostiintelor sa se faca temeinic,rezultand
o baza solida de cunostiinte fundamentale, cu
implicatii practice, care sa permita intelegerea,,
perfectionarea continua si imbunatatirea eficacitatii activitatilor
veterinare.
Adresandu-se,
in primul rand, studentilor din anul I, lucrarea a fost conceputa
tinandu-se cont de programa programa analitica a disciplinei de
Biologie celulara, histologie si embriologie de la Facultatea de
medicina veterinara din Bucuresti, unde autorul este profesor
titular.
Astfel, "Citologia " studiaza ultrastructura si functiile
celulelor, permitand cunoasterea substratului celular al fenomelor
biologice. 'Histologia' studiaza organizarea microscopica a
tesuturilor si organelor ce alcatuiesc organismul animal, iar
'Embriologia' se ocupa cu studiul dezvoltarii ontogenetice,
din momentul producerii
gametilor, al aparitiei oului
(sau zigotului) si pana la formarea intregului organism animal.
Intreaga
lucrare va cuprinde doua volume:
Volumul I - Citologia
si embriologia generala
Volumul II -
Histologia tesuturilor si
organelor, embriologia speciala
Organismul animalelor domestice contine
o serie de substante anorganice (apa, elemente si saruri
minerale, etc.) si organice (proteine,glucide,lipide, enzime, acizi
nucleici, etc.), care nu formeaza solutii la intamplare, ci sunt
organizate cu mare precizie, intracelular sau extracelular, sub forma unor
structuri si ultrastructuri, care indeplinesc diverse functii.La
randul lor, celulele sunt grupate, morfologic si functional in
structuri organizate, tesuturi sau organe, rezultand organismul, ca un tot
unitar.
Celulele care intra in alcatuirea unui organ
realizeaza o structura precisa, asemanatoare la specii
diferite, fenomen cunoscut sub denumirea
de homeostazie structurala.
Pentru realizarea unei structuri coopereaza un mare numar de celule de
tipuri diferite, care comunica intre ele, isi controleaza forma,
marimea, pozitia, orientarea in spatiu si activitatea in
asa fel, incat fiecare celula si structura in ansamblul sau
sa functioneze optim, obtinindu-se efecte maxime cu minimum de
cheltuieli materiale si energetice.
In cunoasterea stiintifica
medicala, functiile pot fi explicate numai dupa intelegerea
structurilor ce constituie substratul lor, mergandu-se pana la nivelul
molecular. A intelege o structura, un ansamblu de fapte observabile optic
sau electronomicroscopic inseamna sa descoperi conexiunile dintre
elementele componente si sa integrezi aceste conexiuni in gandirea proprie (Diculescu - 1987).
In orientarea
generala a continutului lucrarii s-a tinut cont atat de
faptul ca morfologia microscopica se preda in anul I, cat si de
corelatiile sale cu celelalte discipline fundamentale sau clinice,
implicate in asigurarea bazei de date cu care va opera actualul student,
viitorul medic veterinar in activitatea sa practica si
stiintifica.
Intrucat domeniul abordat cunoaste o gigantica explozie informationala, a
trebuit sa selectam din noianul de informatii numai pe acelea
care prezinta o importanta deosebita pentru latura
formativa,, educativa si stiintifica a
invatamantului medical veterinar. Acest fapt limiteaza, de
la inceput orice pretentii asupra cartacterului exhaustiv al lucrarii
si al indicatiilor bibliografice.
Ilustrarea materialului teoretic este realizata prin
scheme executate de autor, preluate
si modificate din lucrarile de specialitate.
Multumesc tuturor celor care mi-au adresat sugestii
si idei ce au dus la imbunatatirea acestui material
didactic, pe care il doresc a fi cat mai util studentiilor.
1.1
Repere istorice in evolutia cunostiintelor despre celule si tesuturi
Descoperirea celulei a fost posibila dupa
inventarea microscopului in anul 1590
de catre doi frati olandezi HANS
si ZACHARIAS JENSSEN. In anul 1665, ROBERT HOOKE, fizician englez a observat cu ajutorul microscopului,
intr-o bucata subtire de pluta, o serie de cavitati mici pe care le-a denumit celule ( de la latinescul cella = camaruta).
Mai tarziu (1674), Antoine. van LEEUWENHOEK descopera mai multe tipuri de celule libere (protiste, globulele rosii) , iar HAM LUDWIG (1677) a descoperit spermatozoizii. Tot in secolul al XVII-lea, MARCELLO MALPIGHI (1628-1694)
a descris pentru prima data capilarele sanguine in peritoneul de
broasca si le-a denumit astfel dupa latinescul 'capillus', care inseamna fir
de par. Totodata, Malpighi a mai descoperit glomerulii renali,
stratul intermediar al epidermului, dar nu si-a dat sema ca celula
este unitatea de baza a lumii vii. Olandezul Regnier de GRAAF
(1641-1673) a descoperit foliculii ovarieni.
La sfarsitul secolului al XVIII-lea, FRANCOIS MARIE XAVIER BICHAT (1771-1802)
a descris si a introdus notiunea de tesut,
individualizand 21 tipuri de tesuturi si descriind in mod magistral
tesutul osos, cartilaginos si muscular.Pentru descoperirile sale este
considerat parintele histologiei.
Un moment crucial in dezvoltarea cunostiintelor
despre celule l-a avut cristalizarea teoriei celulare pe baza cercetatilor
si generalizarilor efectuate de botanistul
MATHIAS JACOB SCHLEIDEN (1804-1880)
si de zoologul THEODOR SCHWANN
(1810-1882). Ei au aratat ca celula nu este goala, ci
contine un 'utricul azotat',
cu caracter reticular, ce se formeaza din citoblastem.
Teoria celulara, elaborata de
ei, sustinea ca: 1- corpul tuturor plantelor si animalelor este
format din celule; 2- celulele au o viata proprie,
caracteristica fiecarei
celule; 3- viata individuala a celulelor este influentata
de viata in ansamblu a organismului.
Mai tarziu, RUDOLPH VIRCHOW (1821-1902) a completat si
imbunatatit teoria celulara, sustinand ca: 1)
'Omnis cellula e cellula'
-toate celulele provin din celule preexistente, negand in acest fel
generatia spontana din citoblastem; 2) 'Omnis vivum ex ovo' - tot ce este viu provine din diviziunea
si cresterea unui ou fecundat, rezultat din contopirea a doua
celule diferite, ovulul si spermatozoidul; 3) In afara celulelor nu
exista viata, iar organismul este o confederatie
de celule.
Microscopul optic modern a aparut in a doua jumatate a secolului a XIX-lea
ca urmare a contributiei unor fizicieni, printre care se disting doi
germani, mecanicul KARL ZEISS
(1816-1888) si profesorul ERNEST
ABBE
Primul microscop
electronic a fost pus la punct de germanii G.A. KAUSCHE si H. RUSKA in 1939, cand au reusit
sa observe si sa fotografieze un virus. In paralel cu
perfectionarea microscopului optic,s-au pus la punct tehnicile de colorare si impregnare cu substante de contrast a componentelor celulare, in acest
domeniu remarcandu-se germanii G.H. HOFMANN,
R. HEIDENHEIM, C.WEIGERT, R. FEULGEN, A. PAPPEMHEIM,
P.EHRLICH, G.MANN, danezii M.G. GRAM,
italianul C. GOLGI, spaniolul RAMON Y.CAJAL, romanii V.BABES, G.LEVADITI, D.VOINOV
si altii.
In secolul XX,
histochimia si histoenzimologia moderna au fost
fondate si ilustrate de personalitati ca: R. FEULGEN, J.VERNE, J.BRACHET, G.G M RI,A. SELIGMAN, A.C.PEARSE si altii.
1.2. Contributii romanesti
la studiul celulei si la dezvoltarea
histologiei
Constituirea unei discipline de histologie de sine
statatoare in invatamantul medical din Romania s-a
realizat in anul 1873, la Iasi de catre profesorul SCULLY si in anul 1881
la Bucuresti de catre
profesorul M. PETRINI-GALATZI, elev
a lui RANVIER, prin separarea
histologiei de anatomie. Inainte de aceste date,insa N. KRETZULESCU (in 1839) si-a
sustinut teza de doctorat privind structura
microscopica a dintilor si cristalinului, iar din 1859 LUDOVIC FIALLA a predat un
curs de Anatomia microscopica si histologie, fiind primul
profesor de histologie la scoala de medicina din Bucuresti, ctitorita
de Carol Davilla.
La Facultatea de Medicina Veterinara din Bucuresti, in
anul 1883,histologia figura printre obiectele de studiu ale anului I,
fiind predata de profesorul C.Gavrilescu pana in anul 1899, iar in continuare de profesorul Ion Athanasiu, in paralel fiziologia animala. Din 1905
pana in 1923, cursul de
histologie a fost predat de profesorul Ion
Dragoi, elev al lui Ion Athanasiu, impreuna cu care a efectuat
cercetari remarcabile privind histofiziologia
tesutului muscular, descriind sarcolema
si tubii ' T '. Intre
anii 1923-1941, profesorul
Dragoi a predat histologia la Facultatea de medicina umana din Cluj. Dupa 1923, la conducerea disciplinei, s-au succedat profesorii: G. NIKITA (intre anii 1923-1928), I.T.NICULESCU
(intre 1928-1932)- un stralucit
elev al lui G. MARINESCU (fondatorul neurohistologiei romanesti), ILIE BADESCU (intre 1932-1848), ILIE DICULESCU (intre 1946-1962), V.Ciurea si GH.BORDA ( intre anii
Profesorul ILIE DICULESCU, absolvent al
Facultatii de medicina veterinara, a condus din anul 1962
disciplina de Citologie si histologie, de la Facultatea de medicina
umana din Bucuresti, impunandu-se ca un creator al scolii de histoenzimologie si citochimie
ultrastructurala. Un merit deosebit in dezvoltarea biologiei celulare
revine savatului de origine romana, GEORGE
EMIL PALADE, laureat al premiului Nobel pentru medicina, considerat
principalul cartograf al celulei.
In prezent, disciplinele de Biologie celulara,
histologie si embriologie sunt predate la Facultatile de
medicina veterinara din tara
de: prof.dr. Corneliu Cotea la Iasi,
prof.dr. Viorel Miclaus la Cluj-Napoca, prof.dr. Mariana Sincai la Timisoara si prof.dr. Nicolae Cornila la Bucuresti,
continuandu-se si dezvoltandu-se traditiile valoroase ale intemeietorilor
si inaintasilor Scolii romanesti de morfologie
microscopica.
1.3.Conceptia
actuala despre citologie Citologia este o ramura a
stiintelor biologice, care studiaza structurile functionale si fenomenele biologice generale comune tuturor celulelor,
evidentiind legile generale de
desfasurare a proceselor vitale la nivelul celular si molecular
de organizare. Manifestandu-se ca o stiinta
integrativa, ea foloseste notiuni de morfologie, biochimie
moleculara si celulara, fiziolgie celulara si
genetica moleculara. Ultimul deceniu se remarca prin
intensificarea studierii nivelului molecular al proceselor celulare. Celula este considerata ,astfel, un microcosmos, in care structurile si functiile
interactioneaza armonios, fiind determinate si reglate genetic, incat sa se realizeze o eficacitate maxima.
Dezvoltarea biologiei celulare si moleculare se
datoreaza pe de o parte perfectionarii tehnicilor de studiere a
celulei, iar pe de alta parte progreselor teoretice (conceptuale).
1.4.1.Tehnicile de studiere a
celulei.
Sunt reprezentate de tehnici de microscopie (optica sau
electronica) si de tehnici de cercetare
( fizice, biochimice, debiologie celulara si moleculara , etc).
1.4.1.
Tehnicile de microscopie optica si electronica
Microscopia optica foloseste fotonii pentru realizarea imaginilor, asigurand mariri de 1500-3000 de ori (in mod obisnuit
de 1400 ori), cu o putere de
rezolutie de 0,2 micrometrii ( ).(Fig.1.1)
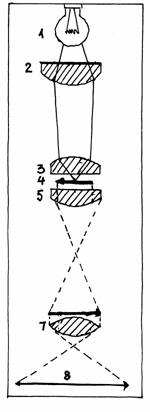
Fig.1.1 Formarea
imaginii in microscopul optic
1.Sursa de lumina; 2.Dispozitiv de uniformizare
si dozare a luminii;3.Condensor;4.Preparat;5.Lentila
obictiv;6.Imagine formata in obiectiv;7.Lentila ocular;8.Imagine
finala.
Microscopia
in contrast de faza, permite examinarea celulelor vii
prin utilizarea: microscoapelor cu
interferenta, pentru aprecierea cantitativa a masei unor componente tisulare; - microscoapelor cu interferenta
diferentiata, pentru studierea suprafetei celulelor.
Microscopia de
fluorescenta evidentiaza moleculele cu fluorescenta naturala (
vitamina A, catecolaminele) sau fluorescenta indusa in imunohistochimie pentru detectarea
reactiei antigen-anticorp.
Microscopia cu ultraviolete foloseste lumina ultravioleta
pentru detectarea unor amino acizi
sau a acizilor nucleici.
Microscopia cu lumina polarizata pentru studierea
formatiunilor cristaline.
Microscopul confocal
Permite obtinerea de imagini
tridimensionale ale structurilor din preparatele histologice.
Principiul de functionare este asemanator
cu cel al microscopului de fluorescenta. Sursa de lumina este
reprezentata de un laser a
carei raza trece printr-o diafragma
cu deschidere punctiforma,
dupa care este reflectata de o oglinda
mobila spre lentila obiectiv. Preparatul este situat la nivelul
distantei focale a obiectivului.
In urma actiunii razei laser
asupra preparatului, acesta emite un fascicul de lumina (eventual
fluorescenta) care este colectat de lentila obiectiv si dirijat spre diafragma
punctiforma a detectorului, situata confocal
(in focarul lentilei obiectiv, opus focarului in care este situat
preparatul), rezultand o imagine
finala, care este captata de un detector
(similar ocularului).
Razele luminoase emise de formatiuni situate in alte planuri nu vor fi captate de
detector, incat se obtine o imagine
foarte'curata', neestompata de imagini ale obiectelor
situate proximal sau distal fata de planul focal. Prin deplasarea in plan vertical a focarului lentilei
obiectiv, se pot obtine imagini
ale unor planuri succesive, ca profunzime, realizand 'sectiuni optice', fara a fi nevoie de o resectionare a
preparatului. Imaginile planurilor succesive sunt stocate in memoria unui
computer, care le 'ansambleaza' obtinand o imagine tridimensionala a preparatului examinat.
Microscopul confocal permite examinatorului sa
obtina 'tomografia'
unui preparat histologic, fiind folosit pentru studirea citoscheletului, a
dispunerii cromozomilor in nucleu,etc. Daca este dotat cu lumina
flurescenta poate obtine
imagini confocale fluorescente. Microscopul
electronic se bazeaza pe utilizarea fluxurilor de electroni pentru realizarea imaginilor,
permitand obtinerea unor grosismente teoretice mai puternice de 10.000 de ori decat in microscopia
optica. Se ajunge la o putere de rezolutie egala cu 0,1 nanometri (nm). Exista doua
tipuri principale de microscoape electronice: - microscopul electronic de transmisie si microscopul electronic de baleaj
(scanning).(Fig.1.2)
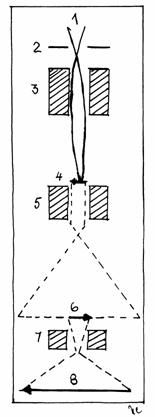
Fig.1.2
Formarea imaginii in microscopul electronic
1-Catod; 2-Anod; 3-Lentila
(bobina)condensator;4-Preparatul; 5-Lentila (bobina) obiectiv; 6-Imaginea
primara; 7-Lentila (bobina) de
proiectie; 8-Imagine finala.
Microscopul
electronic de transmisie (Transmission electron microscope - TEM) poate fi:
- conventional (CTEM), cand
diferenta de potential ajunge pana la 100.000 volti; sau - de inalt
voltaj (HVEM), cand diferenta de potential ajunga la 800.000
volti.
Componentele esentiale ale unui
microscopo electronic de transmisie sunt:
- Catodul sau sursa de electroni, reprezentat de un filament de tungsten
incalzit.
-
Anodul, care relizeaza diferenta de potential.
- Un sistem de electromagneti, care
actioneaza ca niste lentile
condensator, lentile-obiectiv si lentile de proiectie.
-
Un suport pentru preparat sau portobiect, reprezentat de o grila metalica fina pe
care se fixeaza sectiunea de tesut, inclusa in masa
plastica.
-
Un ecran
pe care se capteaza imaginea.
- Un dispozitiv de fotografiere a imaginii.
Puterea de
rezolutie este de 2 Angstromi(Å),
in practica ajungandu-se la 1,2
nanometri.
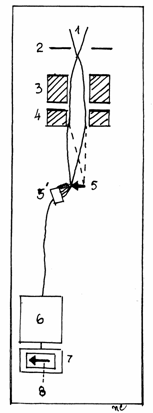
Fig.1.3 Formarea imaginii la
microscopul de baleaj (scanning)
1-Catod; 2-Anod;
3-Lentila (bobina) condensator; 4-Bobina de scanare; 5-Preparat;
5 -Detector de electroni; 6-Amplificator electronic; 7-Ecran de
proiectie;8-Imagine finala.
Ca fixatori se
folosesc glutaralaldehida, pentru constiutienti proteici si tetraoxidul de osmiu
pentru lipide si fosfolipide. Se realizeaza
sectiuni de ordinul nanometrilor cu ajutorul unor ultramicrotoame,
inzestrate cu cutite de diamant. Ultrasectiunile sunt incluse in mase plastice si intise pe grile de cupru. Pentru realizarea contrastului intre diferite
ultrastructuri se folosesc substante precum citratul de plumb si acetatul
de uranil.
La examinare se observa ca structurile cu densitate mare disperseaza (imprastie) electronii,
aparand de culoare inchisa,
in timp ce structurile cu densitate mica, care permit
trecerea electronilor apar, fara a-i devia apar clare.
In cazul microscoapelor
electronice de inalt voltaj (HVEM), diferenta de potential atinge
un milion de volti. Electronii
pot patrunde prin celule intregi
sau preparate de cativa microni (cca. 2
microni). Se obtine o imagine detaliata a organizarii tridimensionale, permitandu-se
vizualizarea texturii celulare interne si a unor detalii de structura,
cum ar fi de exemplu elemntele
citoscheletului.
Microscopul
electronic de scanning (Scanning electron microscope - SEM) se
carcterizeaza prin faptul ca
electronii nu trec prin preparatul studiat, ci matura (sterg)
suprafata acestuia, incat se va obtine o imagine tridimensionala a accidentelor de suprafata.
Puterea de rezolutie este de cca. 100
Angstromi.
Cand electronii folositi de TEM sau SEM
bombardeaza preparatul se produc radiatii
X cu lungimi de unda caracteristice elementelor lovite de electroni.
Prin analiza acestor raze X cu aparate adecvate se pot obtine estimari calitative si
cantitative foarte exacte asupra elementelor cu numarul atomic mai
mare de 12.
1.4.2.Tehnici fizice .biochimice,de
biologie celulara si moleculara
Tehnici fizice
- Fractionarea celulei prin centrifugare diferentiata permite
obtinerea in stare pura a unor componente celulare, precum complexul
Golgi, aparatul mitotic, ribozomi, etc. O contributie deosebita la
perfectionare acestei tehnici a adus-o G.Em.Palade , care a pus la punct centrifugarea
in gel de sucroza.
- Criofracturarea permite despicarea
membranelor celulare inghetate, la nivelul planului median al bistratului
lipidic.
- Autoradiografia
localizeaza materialul radioactiv din tesut.
- Historadiografia realizeaza o microradiografie cu raze
X a unui preparat histologic.
- Difractia razelor X, care a
permis descifrarea structurii
spatiale a proteinelor
si a acizilor nucleici.
- Difractia
si dispersia de neutroni, prin care s-a descifrat organizarea
moleculara a cromatinei.
- Rezonanta
magnetica nucleara de inalta rezolutie, ce se
utilizeaza pentru studierea interactiunilor
moleculare in membranele biologice.
- Rezonanta magnetica in impulsuri,
folosita la studiul permeabilitatii membranelor pentru apa
si ioni.
- Rezonanta electronica de spin,
ce foloseste ca markeri specifici substante cu electroni impari care se atseaza si
evidentiaza molecule de ADN,
de proteine,etc.
- Spectrofluorometria,ce permite studierea
vascozitatii membranelor, difuziunea, rotatia proteinelor.
Tehnici
biochimice
In biologia moleculara se utilizeaza:
- Electroforeza in gel de
poliacrilamida, pentru studiiul proteinelor si acizilor
nucleici.
- Marcarile
bazate pe fotoafinitate a
unor parti din moleculelele integrate in structuri celulare.
- Reconstituiri
de structuri si functii celulare din componentele purificate.
- Imunocitochimia evidentiaza
reactiile dintre antigen si anticorp.
Tehnici complexe
de biologie celulara si moleculara
Sunt reprezentate de:
- Tehnica ADN-ului recombinat.
- Producerea de anticorpi monoclonali cu ajutorul hibridoamelor,
ce a permis descifrarea mecanismelor care regleaza exprimarea genei.
- Sinteza
artificiala a genelor.
- Manipulari de gene, etc.
1.5. Progrese conceptuale.
Progresele teoretice
sau conceptuale au permis
imbogatirea, dezvoltarea si aprofundarea
cunostiintelor de biologie celulara si moleculara.
Sunt reprezentate de:
- Analiza
si sinteza informatiilor obtinute de morfologia
celulara, biochimia si biofizica celulara, fiziologia
celulara,genetica moleculara, imunologia celulara si
moleculara, virusologie,etc. In acest concept se manifesta doua
tendinte opuse: una de acentuare
a sintezei si alta de extindere
a sferei de sinteza.
- Molecularizarea
informatiilor , care consta in descifrarea structurilor
si functiilor pana la
nivel molecular. Acest fapt a dus la schimbarea denumirii acestei ramuri a
stiintelor biologice din 'citologie generala' in 'biologie celulara' si
apoi in 'biologie celulara si moleculara'.
- Integrarea
informatiilor obtinute prin toate metodele si de la toate nivelele intr-un tot unitar, incat biologia celulara si moleculara studiaza in mod
complex si complet aspectele morfofunctionale ale celululelor, observabile la microscopul optic, ultrastructurile
relevate de microscopul electronic, organizarea si fiziologia
moleculara, realizand o imagine
exhaustiva a nivelului celular de organizare a materiei vii.
2.
CELULA CA SISTEM BIOLOGIC
Prin sistem biologic se intelege un ansamblu de elemente (componente) interconectate intr-o formatiune complexa, relativ stabila, formatiune care se
comporta ca un intreg, cu proprietati si functii
distincte cantitativ si calitativ de proprietatile elementelor
componente.
Exista
sisteme lipsite de viata (abiotice),
precum particulele subatomice, atomii, moleculele si sisteme dotate cu
viata (biotice )cum ar fi
celulele, tesuturile, organele, organismul, populatia, biocenoza
si biosfera.
Celula (cellula) este primul sistem
biologic care manifesta cea mai importanta caracteristica a
materiei vii - capacitatea de auto
reproducere. Formele de 'viata
moleculara', identificate pana in prezent, ca 'prionii'
(microorganisme patogene, formate din particule
proteice, fara acizi nucleici)
si virusurile (microorganisme
ce contin acizi nucleici
si proteine) sunt capabile
sa se reproduca numai in interiorul celulei.
Celula, ca sistem, prezinta urmatoarele
caracteristici:
Este un sistem deschis, pentru ca are in
permanenta schimburi de energie si substanta cu mediul
extracelular.
Are un caracter istoric, aparand la un
anumit moment al evulutiei materiei vii si putand sa
dispara, atunci cand materia vie isi va produce o alt
forma mai
eficienta de organizare decat celula.
Are caracter informational, deoarece
receptioneaza, acumuleaza, prelucreaza si transmite
informatii cu ajutorul codului genetic.
Prezinta trei categorii de programe, legate de
capacitatile sale structurale si functionale: a - programul ' pentru sine', ce asigura autoconservarea celulei ( de expl. in
culturile de celule); b - programe ale sistemelor componente, ca
de exemplu programele organitelor sau ale complexelor moleculare; si c - programe superioare care asigura existenta sistemelor
superioare ( tesut, organ, organism).
Echilibrul dinamic, ce se manifesta
printr-o 'pendulare'
continua, fata de o stare
morfofunctionala optima, stabila.
Autoreglarea sau 'feed back'-ul, prin care isi
controleaza procesele interne, in functie de relatiile cu mediul prin mecanisme de tip cibernetic.
Heterogenitatea interna este
insusirea de a fi alcatuita din elemente componente mai mult sau
mai putin diferite ( de expl. mitocondrii, ribozomi, microfilamente,
microtubului,etc.)
Integralitatea, prin care celula ca sistem,
nu se reduce la suma
insusirilor elementelor componente, ci prezinta insusiri noi (structurale si functionale) pe care nu
le au aceste elemente considerate izolat. Aceasta insusire
permite desfasurarea functiilor biologice fundamentale:
metabolismul, reproducerea, adaptarea, mentinerea starii de
stabilitatea diferentiata,etc.
Pe baza acestor caracteristici, conceptia actuala despre celula o defineste ca
fiind: unitatea elementara a lumii
vii, produs al evolutiei,
cu structura complexa, in
relatie de autonomie si echilibru dinamic cu mediul
inconjurator, avand ca proprietati esentiale metabolismul, autoreproducerea, cresterea
si dezvoltarea.
2.1. Arhetipurile celulare
Exista
doua arhetipuri (arhetip =
model primar, dupa G.E.Palade) celulare: procariotele (procariot =prenucleat, fara
nucleu distinct) si eucariotele
(eukariot= cu nucleu distinct).
Procariotele sunt reprezentate, in
principal, de bacterii si de algele albastre verzi (denumite si
cianobacterii). Ele sunt mai mici (1-10 ) decat celulele
eucariote si lipsite de membrane
intracelulare. Organitele celulare sunt putine (ribozomi) sau absente. Nucleul
nu apare distinct, genomul (totalitatea
genelor) nefiind separat de citoplasma. Prezinta un cromozom unic, dispus in
citoplasma si reprezentat de un ADN
circular neconjugat cu proteine.
ARN-ul si proteinele sunt
sintetizate in acelasi compartiment: in citoplasma. Nu au
aparat mitotic, deoarece se inmultesc prin sciziparitate. Isi realizeaza locomotia prin flageli simpli. Au metabolism aerob sau anaerob. Sunt delimitate de o membrana plasmatica, ce
trimite prelungiri endocelulare, denumite mezozomi, dublata la exterior
de un perete celular. Nu prezinta citoschelet, nu au curenti citoplasmatici, iar endocitoza
si exocitoza sunt absente. De
obicei sunt monocelulare.
Eucariotele sunt reprezentate de protiste, fungi, plante si animale.
Au dimesiuni mai mari (5-100 ) decat
procariotele.Prezinta un sistem complicat de membrane intracelulare, care
delimiteaza unele componente intracelulare (nucleul) si unele organite
(mitocondriile, reticulul endoplasmic,complexul Golgi,etc). Au un nucleu distinct, in care este situat genomul, ce cuprinde mai multi cromozomi. Molecula de
ADN este foarte lunga si
conjugata cu proteine. ARN-ul
este sintetizat si procesat in nucleu,
in timp ce proteinele sunt sintetizate in citoplasma. Au citoschelet compus din proteine
filamentoase, curenti citoplasmatici si
desfasoara procese de endocitoza
si exocitoza. Prezinta aparat mitotic.Se divid prin mitoza
si meioza.
2.2 Morfologia generala a celulelor eucariote
Dimensiunile celulelor eucariote
sunt extrem de variate. Ele se
exprima in urmatoarele unitati de masura:
micrometrul, nanometrul si angstromul.
1 micrometru ( m sau ) = 10-3
mm = 10-4 cm = 10-6 m
1 nanometru (nm) =10-3 m= 10-6 mm
= 10-7 cm = 10-9 m
1
angstrom ( )=10-1nm
=10-4 m=10-7mm=10-8 cm =10-10 m
Limita practica de rezolutie a ochiului uman
este de cca. 0,1 mm, a microscopului optic de 0,2 m, iar a celui
electronic de 2 angstromi(
Pentru fiecare categorie de celule, dimensiunile sunt
relativ constante, indiferent de specie sau marimea organului.
Diferentele de greutate a aceluiasi organ la specii diferite sunt
generate de numarul de celule pe care il contin si nu de volumul
celulelor. De exemplu, nefrocitele si celulele musculare cardiace au
dimensiuni apropiate la hominide,ecvine
si canide.
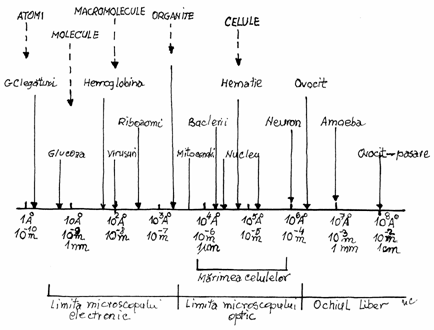
Fig.2.1.Diagrama
unitatilor de lungime
Dimensiunile medii ale
diametrului celulelor se incadreaza
intre 10-30 m, dar in organismul animalelor exista si celule mai mici,
precum: neuronii din stratul molecular al scoartei cerebrale ( de cca.4-4 m), eritrocitele
(5-7 m), precum si celule ce ajung la dimesiuni foarte mari, un exemplu
constituindu-l ovulul ce atinge un diametru de 150-200 m la mamifere,
3,5 cm la pasari si 10 cm la strut. Neuronii motori din
coarnele ventrale ale maduvei spinarii au un corp ce atinge un
diametru de 200 m, iar daca se iau in considerare si prelungirile ajung la o
lungime de 1-1,5 m sau mai mult, in functie de talia animalului. Fibra
musculara striata are un diametru de ordinul micronilor, dar lungimea
este foarte variata, de la 1 la 12 cm.
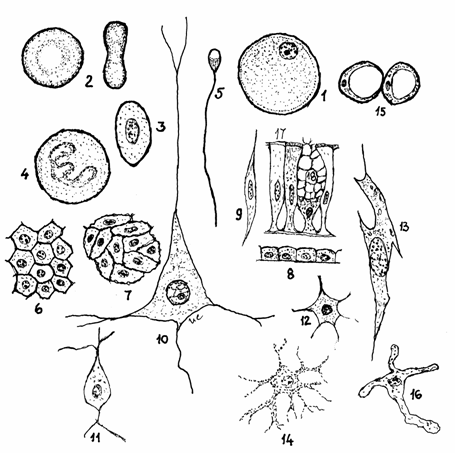
Fig.2.2.
Forma celulelor
1-Ovula; 2-Hematie de
mamifer; 3-Eritrocit de pasare; 4-Neutrofil de mamifer; 5-Spermatozoid;
6-Celule poliedrice;7-Celule pavimentoase; 8-Celule cubice; 9-Leiocit;
10-Neuron piramidal; 11-Neuron piriform; 12-Celula mezenchimala;
13-Fibroblast;14-Melanocit,15-Adipocit alb; 16-Histiocit; 17-Celula
prismatica cu platou striat; 18-Celula caliciforma.
Volumul celulelor variaza in
limite foarte largi de la 200 la15000 micrometri cubi ( ),
mentandu-se constant pentru un anumit tip de celula, independent de
marimea organului, aspect cunoscut sub denumirea de 'legea
constantei volumului'.
Varsta si
functia determina variatii in marimea celulelor. Se
observa o reducere a volumului celular in cursul embriogenezei timpurii,
in cursul imbatranirii si o crestere volumetrica in timpul
dezvoltarii postnatale. Se stabileste un raport nucleo/citoplasmatic
optim, un raport al suprafetei celulei fata de volum si un raport intre metabolism
si volumul celulei.
Masa (greutatea) unei celule este de
aproximativ 10-12 g, iar cea
a unui ovocit uman de 10 -8 g.
Forma celulelor, intial
sferica se modifica in cursul diferentierii si
maturarii celulelor. Forma celulelor este controlata atat de factori
externi ca presiunea si vascozitatea mediului, cat si de factori interni ca activitatea
functionala, varsta, vascozitatea citoplasmei, structura interna
si caracteristicile suprafetei.(Fig.2.2 )
Forma celulelor se adapteaza la functie,
respectandu-se o lege generala a biologiei, conform careia se obtine maximum de randament
functional cu minimum de cheltuieli materiale si energetice.Astfel,celulele
contractile sunt fuziforme sau cilindrice, iar celulele specializate in
conductibilitate prezinta numeroase prelungiri.
Numarul celulelor este extrem
de mare, ajungand la ordinul de milioane de miliarde (1014).
Celulele sanguine reprezinta populatia cea mai numeroasa, reprezentata
de mai multe zeci de miliarde. Numarul hepatocitelor si al neuronilor
este de circa 100 de miliarde, iar cel al nevrogliilor este de aproximativ 10
ori mai mare decat numarul neuronilor.
Durata vietii celulei sau
ciclul celular reprezinta intervalul de la aparitia unei celule
pana la intrarea ei in diviziune. Variaza de la 10-20 minute
pana la 109 minute. Celulele bacteriene traiesc cateva
zeci de minute, celulele epiteliale 1-2 zile, hematiile 127 zile, iar unele
celule musculare si nervoase toata viata individului.
La
om, in fiecare secunda au loc peste 4 milioane de diviziuni celulare,
intr-o zi au loc cca. 350 bilioane diviziuni, iar intr-un an 1014
diviziuni, numar egal practic cu numarul total al celulelor ce
alcatuiesc corpul omului adult.
2.3.
Compartimentarea celulelor eucariote
Celula eucariota prezinta trei componenete
principale: membrana, nucleul si citoplasma.(Fig.2.3)
Membrana poate fi situtata la periferie, realizand pe de o parte delimitarea
celulei de mediul extracelular, sau poate fi dispusa in citoplasma,
unde formeaza un sistem de membrane intracelulare cu o suprafata
deosebit de activa biologic, de 10-20 ori mai mare decat cea a membranei
periferice.
Membranele intracelulare, endomembranele sau
citomembranele difera intre ele prin structura, compozitie
chimica si functie. Ele impart celula in doua
compartimente: compartimentul matriceal sau plasmatic 'P' si
compartimentul intracisternal sau 'E'. Exista si un
compartiment secundar sau de tip 'C', reprezentat de matricea mitocondriala si a
cloroplastelor.
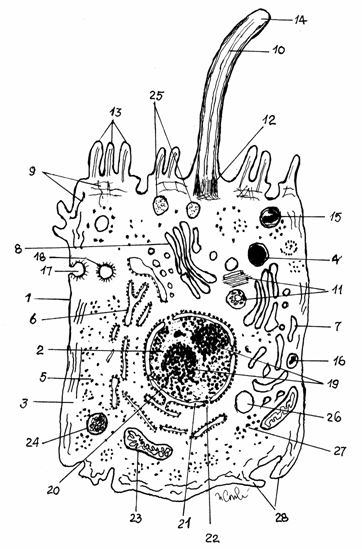
.2.3. Schema unei celule eucariote ideale
1-Membrana
celulara;2-Nucleul; 3-Citoplasma; 4-Lizozom primar; 5-Poliribozomi
liberi; 6-Reticul endoplasmic
rugos; 7-Reticul endoplasmic neted; 8-Complex Golgi; 9- Microfilamente; 10-Microtubuli; 11-Centrioli; 12-Corpuscul
bazal; 13-Microvili; 14-Cil; 15- Fagozom;
16-Peroxizom; 17-Caveola acoperita; 19-Nucleoli;
20-Heterocromatina; 21- Cisterna
perinucleara; 22-Por nuclear; 23-Mitocondrie; 24-Corp rezidual; 25-Granule
de secretie;
26-Picatura de lipide; 27-Granule de glicogen; 28-Pseudopode
Totodata endomembranele delimiteaza nucleul si unele organite celulare
(lizozomii, reticululendoplasmic,complexul Golgi). Exista si organite
nedelimitate de membrane (ribozomii,centriolii, corpusculii bazali,
microfilamentele, filamentele intermediare si microtubulii).
3.
Membranele celulare
Membranele celulare (membrana
celullaris) sunt structuri moleculare complexe care marcheaza limita
celulei sau a unui teritoriu intracelular.
Exista trei categorii principale de membrane
celulare:
1.Membrana plasmatica sau plasmalema,
ce confera individualitate celulei si participa la
interactiunile dintre celule sau la cele cu mediul extracelular.
2.Citomembranele sau endomembranele,
reprezentate de membrana nucleului, membranele reticulului endoplasmic,
membranele complexului Golgi, membranele mitocondriilor, lizozomilor si
peroxizomilor.
3. Membranele speciale reprezentate de membranele tecilor de mielina si de
membranele discurilor din segmentul extern al celulelor vizuale din
retina.
3.1 Organizarea moleculara a membranelor
In evolutia cunostiintelor despre
organizarea moleculara a membranelor se disting mai multe
modele:initial, paucimolecular, unit, mozaicul fluid si modelul
actual.
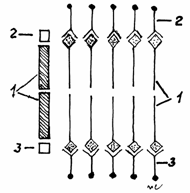
Fig.3.1
Modelul paucimolecular.
1-Bistratul lipidic;
2-Stratul proteic extern; 3-Stratul proteic intern.
Modelul initial, elaborat
de OVERTON in 1895, a sugerat
prezenta lipidelor in membranele celulare, observand ca
solventii organici difuzeaza mai rapid prin membrane decat in
apa. G RTER si GRENDEL (1925) arta ca lipidele din membrane
sunt dispuse in strat dublu (in bistrat), pentru ca suprafata
filmului lipidic din hematii este dubla fata de suprafata
acestora..
Modelul paucimolecular a fost
elaborat in 1930 de DANIELLI si DAVSON, care masurand tensiunea
superficiala a membranelor celulare, au gasit aproximativ o dina/cm patrat, fata de
5 dine cat reiesea din calcule. Asfel, au dedus ca stratul lipidic
dublu (bistratul lipidic) este tapetat pe ambele fete de straturi de
proteine, care reduc tensiunea superficiala. Totodata au demonstrat
experimental acest fapt, adaugand
proteine, ce au produs reducerea
tensiunii superficiale. In 1935, DANIELLI si DAVSON, au elaborat modelul
paucimolecular (paucus = putin numeros), dupa care membranele
celulare au un centru lipoid 'lamina
intermedia', format din fosfolipide, dispuse in strat dublu, cu
capetele polare orientate spre exterior si tapetate cu proteine, care formeaza o lamina externa si alta interna.(Fig.3.1)
Modelul
'UNIT' a fost elaborat de ROBERTSON, intre 1958-1960,
dupa cercetari efectuatte la microscopul electronic ( cu grosisment
de 100.000), efectuate pe membrana tecii de mielina. Dupa acest
model, membranele celulare au o ultrastructura trilaminara,
prezentand doua benzi dense (de 2,5 nm), ce delimiteaza intre ele o
banda clara ( de 3 nm), formata dintr-un ax lipidic hidrofob.
Acest model confirma modele anterioare si le
dezvolta, adaugand doua noi concepte: - universalitatea
bistratului lipidic si - asimetria chimica, dupa care glucidele sunt atasate
numai pe fata externa, lipsind pe fata interna. Modelul
este incomplet pentru ca ignora dinamismul moleculelor
componente.(Fig.3.2).
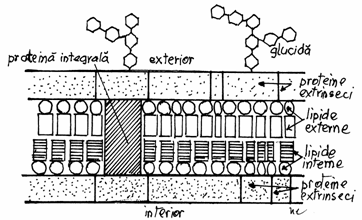
Fig.3.2
Organizarea moleculara a membranei periferice
Modelul'mozaicului fluid',
elaborat de SINGER si NICOLSON (1972) sustine ca moleculele
componente (lipide si protide) difuzeaza in mod intamplator in
bistratul lipidic.
Cercetarile ulterioare
au aratat ca miscarile moleculelor sunt controlate de
citoschelet si sunt limitate la anumite portiuni ale suprafetei
membranei. Exista si zone ale
fetei externe a bistratului lipidic neacoperite de proteine.
Modelul actual sustine
ca:- bistratul lipidic este asimetric, fluid si reprezinta axul
intregului edificiu molecular al membranelor; - pe fetele hidrofile,
proteinele sunt distribuite asimetric, in aranjamente caracteristice.
3.2.
Ultrastructura membranei periferice
Membrana periferica are trei componente
ultrastructurale: plasmalema, glicolema,citoscheletul membranei.
3.2.1.
Plasmalema
Avand o grosime de cca. 7,5 nm, plasmalema (plasmalemma) reprezinta structura de baza a
tuturor membranelor celulare ( periferice sau endomembrane), fiind
alcatuita din bistratul lipidic asociat cu proteine.
La un grosisment
redus (de 10.000) apare ca o linie
unica elecronodensa. La o marire de 100.000 ori, daca
planul sectiunii a cazut perpendicular pe membrana, plasmalema
prezinta un ascect caracteristic, trilaminar, cu doua benzi dense (de
2,5 nm), separate de o banda clara ( de 3 nm). Endomembranele au ,in
general, acelasi aspect electronomicroscopic, respectand modelul unit.
Banda luminoasa, clara (lamina
intermedia) corespunde radicalilor acizilor grasi hidrofobi din
fosfolipidele bistratului, iar benzile
dense (lamina externa et interna)
cuprind gruparile polare hidrofile ale fosfolipidelor si partial
proteinele asociate membranei.(Fig.3.3)
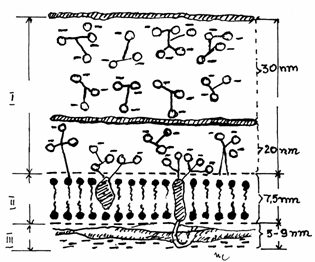
Fig.3.3 Componentele membranei celulare periferice
I-Glicolema; II-Plasmalema;
III-Citoscheletul membranei
Compozitia chimica a plasmalemei cuprinde:
lipide complexe, proteine si glucide.
LIPIDELE sunt continute in bistratul lipidic,
fiind reprezentate de fosfolipide (70%),
colesterol (25%) si glicolipide (5%).
Fosfolipidele
si glicolipidele sunt molecule
complexe cu caracter amfofil (=amfipatice=bimodale), prezentand un
'cap' hidrofil (sau grupare polara) si o
'coada'hidrofoba, ce cuprinde fragmente de acizi grasi
esterificati.Ele au tendinta de a se organiza in pelicule sau
micelii, deoarece cozile trebuie sa se dispuna in afara contactului
cu apa.Mentinerea moleculelor in pelicule sau micelii se realizeaza
prin legaturi hidrofobe ce nu implica cheltuieli de energie.
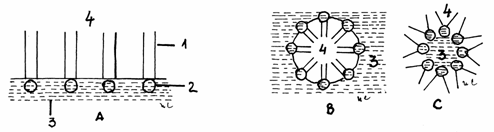
Fig.3.4.
Tendinta de organizare a moleculelor de fosfolipide
A-Monostrat;
B,C- Micelii
1-Laturi de
acizi grasi; 2-Grup polar; 3-Apa; 4-Aer
In cazul fosfolipidelor si glicolipidelor din
membrana, forma cea mai stabila a miceliilor este reprezentata
de stratul lipidic dublu in care gruparile polare sunt dispuse la exterior
in contact cu apa, iar lantul de acizi grasi se afla la
interior. Asfel, se formeaza un bistrat sau continuum lipidic.
Fosfolipidele din plasmalema sunt reprezentate de
fosfogliceride (ca de exemplu: fosfatidilcolina, fosfatidiletanolamina,
fosfatidilserina, fosfatidilinozitolul) si de sfingolipide ( dintre care
cea mai raspadita este sfingomielina. Molecula de
fosfatidilcolina, denumita si lecitina, are aspectul unui
'cui', in care 'capul' (alcatuit din radicali de
colina si fosfat) este hidrofil, poarta sarcini electrice
si este numit grup polar. Resturile de acizi grasi sunt hidrofobe
si formeaza segmentul apolar al moleculei.
Glicolipidele
ocupa 5% din totalul lipidelor si pot fi simple ( de expl.
cerebrozidele) sau complexe (de expl.gangliozidele) Gangliozidele contin
unul sau mai multe resturi de acid sialic (numit si acid
N-acetilneuraminic sau NANA) care le confera sarcina electrica
negativa.
Incarcatura electrica negativa a
suprafetelor celulare este conferita in mare parte de gruparile
polare ale fosfolipidelor, ale glicoproteinelor sulfatate si indeosebi de
radicalii carboxilici ai acidului sialic.
Colesterolul (25%) se
gaseste in proportie mai mare in membranele la care
predomina functia de bariera, dar scade in membranele intracelulare.
Exista o asimetrie in dispunerea lipidelor in
bistratul membranei eritrocitare. Astfel,glicolipidele se gasesc numai in
stratul extern, la nivelul caruia este abundent si colesterolul. In
stratul intern al bistratului lipidic sunt mai frecventi acizii grasi
nesaturati, in timp ce in stratul extern se intalnesc in proportie
mai mare acizii grasi saturati
cu lanturi lungi.Totodata lipidele sunt repartizate neuniform pe
suprafata membranei celulare, putandu-se grupa in 'petice' sau 'calote', cu implicatii
functionale speciale, indeplinind rol de receptori.
Fluiditatea
lipidelor. La temperatura corpului, toate lipidele din membrana se
gasesc in stare fluida. Ele pot executa doua tipuri de miscari: 1-
miscari in interiorul unei molecule, precum miscari ale
atomilor de carbon in lanturile de acizi grasi si in gruparea
polara; 2-miscari ale intregii molecule, ce pot fi: - de
translatie sau de difuziune laterala; - de rotatie in jurul axei
longitudinale a moleculei si c- miscari de difuziune
transversala (flip-flop).(Fig.3.5)
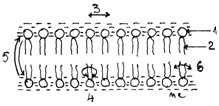
Fig.3.5.Miscarile
lipidelor in bistrat
1-Gruparea hidrofila;
2-Gruparea hidrofoba; 3-Difuziune laterala; 4-Rotatie;
5-Flip-flop;6-Flexie.
Moleculele de lipide se deplaseaza mai usor
prin miscari laterale in acelasi strat si trec foarte
greudintr-un strat in altul.
Unele lipide (expl. cardiolipina) sunt capabile, in prezenta ionilor de calciu, sa
se detaseze din straturile lipidice si sa se dispuna in
forma hexagonala, in cilindri de lipide, care delimiteaza canale, ce permit schimburi selective prin
bistratul lipidic.(Fig.3.6).
PROTEINELE plasmalemei formeaza
doua straturi asimetrice, ce tapeteaza bistratul lipidic sau pot
traversa bistratul lipidic, unde executa miscari de
translatie (difuziune laterala) si de rotatie. Prin
electroforeza in gel de poliacrilamida au fost identificate 15
proteine majore, cu greutati moleculare intre 15.000-250.000 daltoni.
Trei din aceste proteine (spectrina, glicoforina si proteina benzii 3)
ocupa mai mult de 60% din greutatea proteinelor membranare.
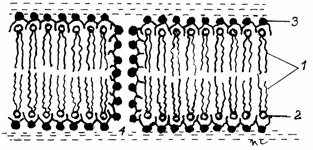
Fig.3.6.Formarea
porilor hidrofili
1-Lipide - grupari hihrofobe; 2-Lipide -
grupari hidrofile; 3-Proteine;4-Por hidrofil
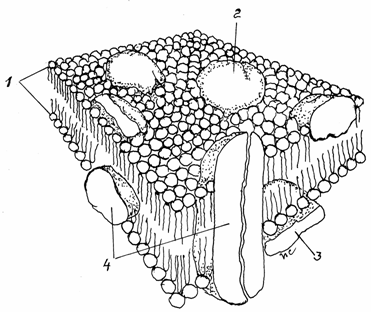
Fig.3.7. Dispunerea proteinelor in plasmalema
1-Bistratul
lipidic; 2- Proteine extrinseci externe;
3-Proteine
extrinseci interne; 4-Proteine intriseci transmembranare
Prin metode de cogelare-fractionare si
fractionare-purificare s-au identificat doua grupuri de proteine:
-a) proteine transmembranare (intriseci sau integrale)
cuprinse in bistratul lipidic ca niste cuburi de gheata, legate
hidrofil si hidrofob de lipide. Se extrag foarte greu si servesc ca dispozitive de
receptie-transductie.
-b) proteine membranare periferice (sau extrinseci)
situate de o parte si de alta a bistratului lipidic, de care se
ataseaza prin slabe legaturi hidrofile.(Fig.3.7)
Proteinele extrinseci sunt
distribuite pe o fata sau alta a bistratului lipidic si si
se leaga de capatele hidrofile ale lipidelor. Sunt mai numeroase pe
fata interna unde participa la realizarea citoscheletului
membranei celulare.
Proteinele de pe fata externa a bistratului
lipidic sunt glicoproteine si se prelungesc pe distante mai scurte
sau mai lungi in glicolema, atasand pe lantul lor polipeptidic fragmente de
oligozaharide. Compozitia, ordinea moleculelor si dispunerea
lanturilor de de oligozaharide fata de proteinele
transmembranare determina specificitatea functionala si
identitatea fiecarui tip de celule.
3.2.2.
Glicolema sau glicocalixul
Glucidele plasmalemei sunt reprezentate de hexoze,
hexozamine si acid sialic. Fragmentale de acid sialic ocupa
intotdeuna extremitatile periferice ale lanturilor de
oligozaharide si confera sarcina electrica negativa
suprafetei celulelor eucariote.(Fig.3.8)
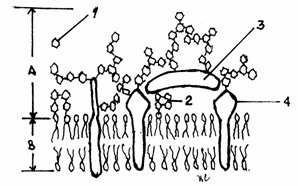
Fig.3.8.
Organizarea glicocalixului
A-Glicocalix;
B-Bistratul lipidic.
1-Rest glucidic;
2-Glicolipida; 3-Glicoproteina adsorbita; 4-Proteina
transmembranara
Lanturile de oligozaharide sunt ancorate pe versantul
extern al plasmalemei, formand un invelis al suprafetei celulare,
numit glicocalix (invelis dulce) sau glicolema. Nu exista o
delimitare neta intre glicocalix si substanta vie din
spatiul extracelular, cunoscuta sub numele de matrice
extracelulara.
Glicolema sau glicocalixul (glicocalyx) prezinta o componenta externa, de natura
glicoproteica, cu grosimea medie de 50 nm, a membranei celulare
periferice. A inceput sa fie investigat abia din 1963, cand BENNET i-a
cercetat structura si functia, implicandu-l in legarea selectiva
a ionilor de calciu si potasiu.
Glicocalixul prezinta doua zone: - o zona
interna, denumita si invelis de suprafata, mai
putin densa, cu grosimea de 20 nm si o zona externa
mai densa, cu grosimea de 30 nm.
Grosimea glicocalixului apare variata, fiind: -mult
redusa la celulele in culturi; - subtiat pana la 20-30 nm, cand
ocupa spatiul dintre celulele invecinate; - crescut, pana la 50 nm la
celulele libere sau pe polul apical al celulelor intestinale.
Din punct de vedere chimic, glicocalixul este
alcatuit dintr-o tesatura delicata de de lanturi
proteice, pe care sunt ancorate fragmente de glucide. Dintre monozaharide, in
glicoproteinele si glicolipidele de la nivelul membranelor si din
glicocalix, cele mai importante sunt: galactoza, galactozamina, manoza,fucoza, glucoza, glucozamina si
acidul sialic.
Raportul strans, de continuitate, dintre glicolema
si plasmalema se datoreaza glicolipidelor si
glicoproteinelor componente, invelisul de suprafata devenand parte integranta a
membranei celulare.
Glicocalixul confera individualitate biochimica
celulei, comportandu-se ca o 'carte de identitate a celulei',
datorita numeroaselor variante moleculare pe care le realizeaza cu
ajutorul monozaharidelor componenete. Intre moleculele membranei,
oligozaharidele induc o variabilitate periferica pronuntata, in
contrast cu varibilitatea redusa a lantului polipeptidc. Astfel, trei
aminoacizi diferiti pot da nastere la sase tripeptide, in timp
ce trei monozaharide pot genera 1056 trizaharide diferite.
Functiile glicocalixului sunt :
Participa,
alaturi de matricea extracelulara, la realizarea
adezivitatii intercelulare;
Asigura
specificitatea si individualitatea biochimica tipului de
celule.Astfel grupele sanguine sunt determinate de gruparile glucidice
terminale aflate pe glicoproteinele si glicolipidele din membrana
eritrocitelor.
Contine
gruparile glucidice ale proteinelor-receptori din membrana, dar
si ale unor proteine implicate in fenomenele de transport.
Depoziteaza
ionii de calciu (Ca2+)datorita sarcinilor elctrice negative
si intervine in controlul schimburilor ionice transmembranare.
Participa
la transmiterea informatiei despre celula.Astfel, unele celule din ficat si din splina
recunosc cu ajutorul receptorilor de membrana glicoproteinele si
glicolipidele de pe suprafata eritrocitelor uzate, care au pierdut acidul
sialic si prezinta descoperita galactoza, ce ocupa penultimul loc din lant.
Galactoza este recunoscuta de receptorii macrofagelor din ficat (celulele
Kupffer), incat hematiile uzate sau imbatranite sunt imobilizate,
fagocitate si lizate.
3.2.3.
Citoscheletul
Citoscheletul (cytoscheleton)
membranei celulare este o retea cu grosimea de 5-9 nm, de proteine
extrinseci, situate pe fata
interna a membranei periferice. Citosheletul este ancorat de
plasmalema prin intermediul capatului intern al proteinelor
transmembranare, iar spre interiorul celulei se continua cu citoscheletul
din matricea citoplasmatica.
La microscopul electronic,
citoscheletul membranei apare ca o retea de microfilamente proteice
orientate neregulat, in nodurile retelei fiind prezente proteine
globulare.
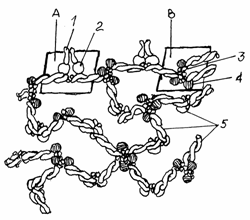
Fig.3.9.Schema citoscheletului
membranei
A-Complexul spectrina,
anchirina, proteina integrala;
B-Complexul spectrina, actina,proteina
benzii 4-1;
1-Proteina transmembranara;
2-Anchirina; 3-Filamente scurte de actina; 4-Proteina benzii 4-1; 5-Spectrina.
Datele referitoare la citoscheletul mebranei au fost
obtinute in urma cercetarilor efectuate pe membranele globulelor
rosii, ulterior fiind considerate valabile si pentru membranele altor tipuri de celule.(Fig.3.9)
In membrana hematiilor, citoscheletul inglobeaza
circa 60% din masa proteinelor intregii membrane. Principalele tipuri de
proteine citoscheletale sunt: spectrina,anchirina, actina,glicoforina si o
proteina care in cursul electroforezei in gel de poliacrilamida
migreaza in banda 4-1. Mai sunt prezente si alte proteine neidentificate, ce au mase
moleculare diferite. Prin extragerea spectrinei si actinei, citoscheletul
membranei se dezintegreaza, piezandu-si forma si structura.
Spectrina (sau
tectina) este proteina care predomina in citoschelet. Apare sub forma a
doi dimeri (unul de 240.000daltoni
si altul de 220.000 daltoni). Dimerii de spectrina au o forma de
bastonas cu o lungime de 100 nm si o grosime de 5 nm. Ei se pot lega
cap la cap formand tetrameri, hexameri, octameri. Moleculele solitare de
spectrina sunt foarte flexibile.
Anchirina (nectina
sau sindeina) este un polipeptid de 200.000 daltoni, cu forma sferica
sau piramidala. Intr-o hematie de la om exista cca. 120.000 de molecule de anchirina, intr-un neutrofil
- 600.000, intr-un trombocit - 3.000,
iar intr-un fibroblast - 30.000. Leaga spectrina la capatul citoplasmatic al proteinei
benzii 4,1.
Actina se gaseste sub forma de mici
fragmente de actina fibrilara, ce contin 13 monomeri. Lungimea
fragmentelor de actina variaza in functie de numarul moleculelor de spectrina pe care le
leaga prin capetele lor, fapt ce poate explica plasticitatea
citoscheletului membranei, in vivo si aspectul lax al acestuia la
microscopul electronic.
Proteina benzii
4-1 are aspect globular, cu diametru de 6 nm.
Laturile retelei citoscheletului membranar sunt
alcatuite din dimeri de spectrina, iar nodurile retelei
(complexele jonctionale) sunt formate din doua tipuri de complexe
moleculare: - un complex ce contine spectrina, anchirina si
o proteina transmembranara, ce realizeaza legatura cu
membrana plasmatica; - si un alt complex format din spectrina,
actina, proteina benzii 4-1 si adducina, prezent in nodurile
propiu zise ale retelei. Proteinele citoscheletului membranei,
impreuna cu lipidele si proteinele de pe fata interna a
plasmalemei prezinta numerosi radicali hidrofili electronegativi, incat
incarcatura electrica negativa este mai puternica
decat pe fata externa.
Totodata, in citoscheletul membranei, se gasesc
unele proteine cu patru locusuri de legare a calciului, denumite calmoduline, ce joaca un rol important in reglarea proceselor intra
celulare. Legaturile dintre diferitale tipuri de proteine din citoschelet
sunt realizate prin procese de fosforilare in care intervine o proteina reglatoare, gelsolina, dependenta de calciu.
Citoscheletul membranei confera: -1) elasticitate,
prin dispunerea in retea a proteinelor; si -2) rezistenta,
prin complexele proteice de la nivelul nodurilor.
3.3.Diferentieri de suprafata ale
membranei celulare periferice.
Sunt neregularitati ale citoplasmei periferice,
cu caracter tranzitoriu sau permanent, delimitate de membrana.
Diferentierile tranzitorii apar
si dispar in raport cu anumite momente functionale ale celulei, avand
aspectul unor infundaturi (invaginatio
cellularis) ale plasmalemei ( in endocitoza), sau al unor expansiuni (processus celularis), cum ar fi
pseudopodele sau valurile ondulante, prin care se realizeaza
deplasarea celulelor.
Pseudopodele (processus amoeboideus) sunt expansiuni cu forma de conuri (processus filiformis) sau degete (processus digitiformis), cu ajutorul
carora leucocitele adera de suporturi, deplasandu-se in timpul
diapedezei. Ele contin organite celulare, precum mitocondrii, ribozomi,
lizozomi.
Valurile si membranele ondulante (processus lamellosus) sunt expansiuni lamelare, foarte
mobile, ce se formeaza in mediul lichid. Nu contin organite si
nu adera la suporturi. Apar in urma unor modificari ale tensiunii
superficiale si se intalnesc la
histiocitele implicate in fagocitoza.
Expansiunile permanente sunt
reprezentate de microvilozitati, cili si flagel.
Microvilozitatile (microvillus) sunt expansiuni cilindrice, delimitate de membrana polului apical (apex cellularis).Ele intervin in
procesele de absorbtie prin marirea suprafetei de contact a
celulei cu substantele ce vor fi absorbite si prin concentrarea unui
numar mare de receptori. Pot fi
dispuse grupat sau solitar. Microvilozitatile solitare sunt inegale,
cu forme si dimensiuni variabile, distantate unele de altele, formand
marginea in perie ( la nefrocite, din
tubii contorti ai nefronului).
Platoul striat cuprinde numeroase microvilozitati
(300-3000/enterocit) uniforme ca lungime (0,6-0,8 m) si
diametru (200 nm), imbracate in glicocalix. Plasmalema
microvilozitatilor contine ATP-aza si un numar
mare de transportori. Citoplasma microvilozitatilor cuprinde in zona
centrala 10-40 microfilamente de actina, dispuse paralel cu axul lung
al microvilului.(Fig.3.11)
Microfilamentele de actina se
insera cu un capat pe plasmalema polului apical al microvilului, iar
la polul bazal al acestuia se termina intr-o retea perpendiculara
de filamente de actina si miozina, denumita buton terminal,
ce are rolul de a mentine
pozitia microvilului. Manunchiul de microfilamente din microvil este
legat de plasmalema, din loc in loc,printr-o proteina, denumita fimbrina. Aceste legaturi formeaza o scara, in
spirala, de-a lungul microvilului.
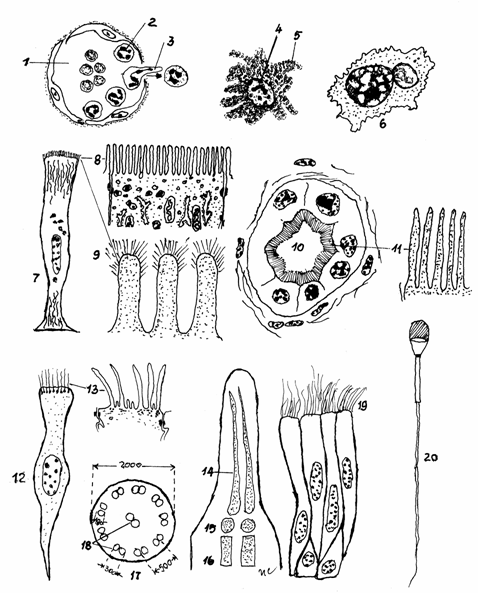
Fig.3.10.
Diferenteri ale membranei celulare
1-Capilar sanguin;
2-Neutrofile; 3-Pseudopod; 4-Nucleu de histiocit; 5-Valuri ondulante; 6-Macrofag cu valuri;
7-Enterocit;8 -Microvili; 9-Glicocalix; 10-Lumenul nefronului; 11- Margine in perie; 12-Celula
prismatica ciliata; 13-Cili; 14-Axonema; 15_Corpuscul bazal; 16-Radacina cilului;
17-Sectiune transversala prin tija cilului; 18-microtubuli centrali
si periferici; 19-Stereocili pe
celule epiteliale din epididim; 20-Flagelul spermatozoidului
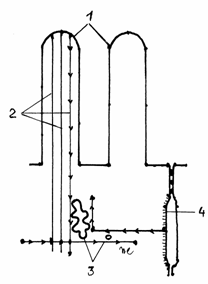
Fig.3.11.Dispunerea microfilamentelor
in microvilul intestinal
1-Microvili;2-Microfilamente
de actina; 3-Microfilamente
de miozina; 4-Zonula adherens.
Cilii (cilium) pot fi mobili (sau vibratili) si rigizi (sau
stereocili).
Cilii mobili (kinetocilii)
sunt prezenti la polul apical al celulelor epiteliale din mucoasa
aparatului respirator sau a oviductului. Au o lungime de 5-15 m, o grosime de
0,2 m si sunt formati din trei portiuni: - tija sau
portiunea libera, -corpusculul bazal si radacina.
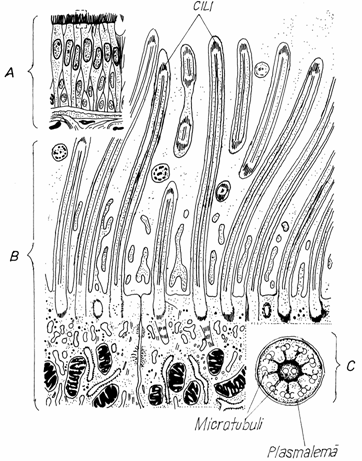
Fig.3.12.Cili-
aspect, structura si ultrastructura
A-Epiteliu pseudostratificat
prismatic cilat; B-Celule prismatice ciliate,polul apical;C-Sectiune
transversala prin cil.
Tija (pars extracellularis) contine o matrice electrono -densa, plasata la periferie
si un complex filamentos axial (sau axonema)(filamentum axiale), format din 10 perechi de tubuli longitudinali:
-o pereche centrala (microtubulus
centralis) si - noua perechi periferice (diplomicrotubulus periphericus), dispuse in jurul perechii
centrale.
Corpusculul bazal (corpusculum
basale) are un aspect cilindric si cuprinde in
strucutra sa noua dublete sau triplete (triplomicrotubulus)
periferice de microtubuli, lipsandu-i perechea centrala. Coordoneaza miscarea cililor.(Fig.3.12)
Radacina (radix basalis) cilului este
formata din dubletele sau tripletele periferice ale corpusculului bazal
care ajung in citoplasma polului apical al celulei ciliate. Are rolul
de a ancora cilul in citoplasma, prezinta
proprietati contractile si participa la conducerea stimulilor receptionati de
portiunea libera.
Stereocilii
sunt cili rigizi, in structura
carora lipseste perechea de microtubuli centrali, avand numai cele
noua dublete periferice. Se intalnesc pe polul apical al celulelor din
epiteliul epididimului.
Flagelul (flagellum) are in axonema o pereche centrala
si noua perechi periferice de microtubuli. Este prezent in
alcatuirea spermatozoidului.
3.5. Jonctiunile celulare
Jonctiunile celulare (junctio intercellulares) sunt structuri
stabile prin care plasmalemele celulelor interactioneaza specific. In
raport cu functiile lor , se disting trei categorii de jonctiuni: -de adezivitate, - impermeabile
si -de comunicare.(Fig.3.13)
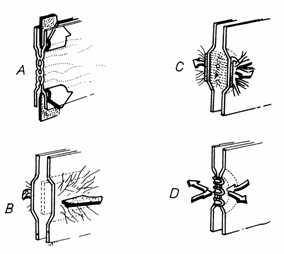
Fig.3.13.Tipuri de
jonctiuni celulare
A-Jonctiuni impermeabile
- zonula occludens; B-Desmozomi in banda - zonula adherens; C-Desmozomi in pata - macula adherens;
D-Jonctiuni permeabile -gap.
In afara acestor tipuri principale se mai
descriu jonctiuni simple si
complexe jonctionale (jonctio
intercellularis complex). In
cazul jonctiunilor simple (junctio
intercelluaris simplex), intre plasmaleme exista un spatiu de cel
putin 30 nm prin care pot trece toate tipurile de molecule existente in
mediul intercelular. Dupa aspectul lor, jonctiunile simple sunt de
trei tipuri: - spatii intercelulare, - jonctiuni intercelulare
denticulate (junctio intercellularis
denticulata), si - jonctiuni intercelulare digitiforme (junctio intercelluaris digitiformes).
3.3.1.Jonctiunile
de adezivitate sau dezmozomii.
Desmozomii (desmosoma)
sunt jonctiuni ce confera o mare rezistenta mecanica
unor tesuturi si organe, solicitate in acest sens . Se prezinta sub trei forme, toate
intalnite in tesuturile epiteliale: 1-desmozomi in pata; 2-desmozomi
in banda; 3-hemidesmozomi.
Desmozomii in pata (macula adherens) .Se mai numesc si desmozomi in 'spot', in
'nit', sau macula adherens
(macula = pata, in limba latina). Sunt prezenti mai ales in
tesuturile epiteliale de acoperire. In alcatuirea lor intra
urmatoarele componente: 1- plasmalemele adiacente, dispuse paralel, la o
distanta de 25-30 nm; 2-un material proteic, intercelular dens la
fluxul de electroni, cu aspect filamentos, bisectat de o densificare
centrala,bogata in glucide si calciu; 3-densificari
intracelulare, in forma de disc, atasate de plasmalele
jonctionate; 4-dispozitive de legatura sau linkeri, reprezentati de microfilamente, ce se
detaseaza din materialul dens intercelular, strabat plasmalemele
si apoi discurile intracelulare, pentru a se ancora la microfilamentele
citoscheletului; 5-elemente citoscheletale, respectiv microfilamente de
actina sau filamente intermediare de cheratina, care se numesc
tonfilamente.(Fig.3.14)
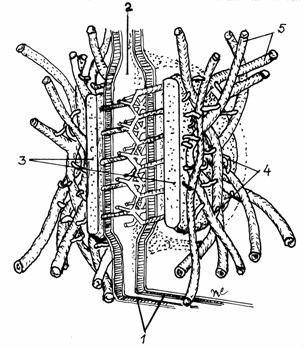
Fig.3.14 Desmozom
in pata-schema
1-Plasmalemele adiacente; 2-
Spatiu si material intercelular; 3-Discuri intracelulare; 4-Linkeri;
5-Tonofilamente.
Desmozomii in
banda (zonula adherens) sau in panglica (zonula=panglica, in latina) sunt
prezenti la polul apical al
celulelor epiteliale si la nivelul segmentelor transversale ale discurilor
intercalare cardiace.
Prezinta o
structura asemanatoare cu cea a desmozomilor in pata, cu
urmatoarele deosebiri: -spatiul intercelular, de 15-25 nm este mai
sarac in material electronodens; -densificarile intracelulare, de pe fata
interna a plasmalemelor jonctionate, nu au forma de disc, ci de
banda sau panglica. (Fig.3.15)
Hemidesmozomii (sau semidesmozomii) (hemidesmosoma) reprezinta o varianta a desmozomilor in
pata, prin care se leaga celulele epiteliale de membranele bazale.
Prezinta numai jumatate din structura desmozomului in pata, in
sensul ca, pe frotul citoplasmatic al plasmalemei exista un disc,
care se leaga de membrana bazala prin filamente.
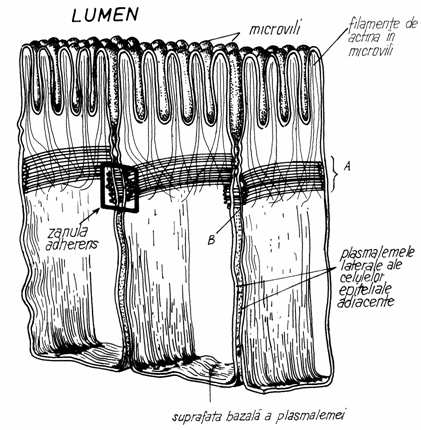
Fig.3.15.
Schema desmozomilor in banda (zonula adherens)
A-Densificari
intracelulare in banda; b-Spatiul intracelular.
3.3.2.Jonctiunile
impermeabile ( sau
jonctiunile stranse).
Sunt dispuse in panglica, fapt pentru care se mai
numesc zonula occludens. Se caracterizeaza prin
obliterarea spatiului intercelular, deoarece membranele adiacente se
apropie complet sau se sudeaza pentru a forma structuri pentalaminate sau
heptalaminate. La realizarea acestor jonctiuni participa proteine transmembranare, care se dispun in
siruri gemene pentru a construi dispozitive ce se conecteaza 'in
fermoar' pe fetele externe ale membranelor adiacente. Pe fetele
interne , proteinele transmembranare sunt 'ancorate' prin intermediul
unor microfilamente la citoscheletul matricei.
Aceste jonctiuni impiedica scurgerea fluidelor
printre celule, inclusiv a micromoleculelor, care sunt obligate sa treaca prin celula. Sunt intalnite
la portiunile apicale ale celulelor care delimiteaza lumenul
intestinului. Astfel, glucoza este pompata activ din lumenul intestinal in
celula prin
suprafata polului apical, dupa care trece in sange prin difuziune
facilitata mediata de transportorii situati in zonele
bazo-laterale ale plasmalemei, pe care o polarizeaza functional
intr-un domeniu
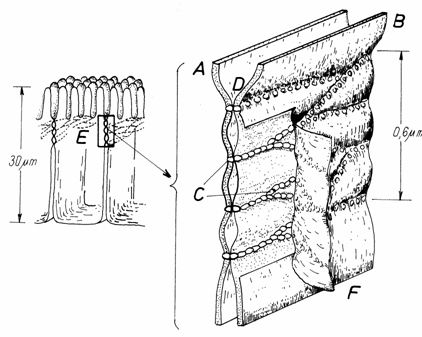
Fig.3.16.Schema
jonctiunilor impermeabile
A-,B-Plasmalemele adiacente;
C-Proteine transmembranare; D-Spatiu intercelular;
E-Enterocit;
F-Citoscheletul matricei.
apical si altul
latero-bazal.Totodata, jonctiunile stranse impiedica
retrodifuziunea glucozei din
spatiul intercelular in lumenul intestinului.(Fig.3.16).
Jonctiunile stranse au aspecte diferite, in functie de
tesutul epitelial in care sunt prezente. Astfel, jonctiunile stranse
de la polul apical al celulelor din tubul contort proximal al nefronului au o
rezistenta relativ scazuta, fiind formate numai din unul sau
doua siruri de proteine transmembranare. La nivelul celulelor
epiteliale din mucoasa vezicii urinare, jonctiunile stranse contin
sase sau mai multe siruri de proteine transmembranare, incat se
realizeaza o jonctiune puternica, extrem de greu de traversat.
La nivelul barierei hematoencefalice, jonctiunile stranse existente intre
celulele endoteliale ale vaselor din encefal realizeaza o protectie
eficienta pentru tesutul nervos, permitand insa trecerea
glucozei (principala sursa de energie) din sange in creier si
trecerea, in sens invers a bioxidului de carbon, rezultat din respiratia
celulelor nervoase.
Jonctiunile celulare stranse se caracterizeaza
prin: -flexibilitate si siguranta; - formarea unor bariere
chimice si fizice intercelulare; - conferirea unei polaritati a
celulelor angajate in jonctiuni; -aparitia de timpuriu, intre
celulele embrionare, initial sub forma de macule, care se extind
treptat, luand forma de zonule.
Jonctiunile focale reprezinta o varianta a
jonctiunilor stranse, in care membrana celulei se apropie de substratul de
adezivitate pana la 10-15 nm. Pe fata extracelulara a membranei
sunt prezente filamente glicoproteice de fibronectina, iar pe fata
intracelulara se aglomereaza fascicule de microfilamente de actina si
vinculina, care se leaga de plasmalema prin intermediul unor
densificari adiacente membranei. Intre filamentele de fibronectina
si microfilamentele de actina si vinculina se
realizeaza dispozitive transmembranare de legatura (de 8-20 nm),
denumite fibronexuri.
3.3.3.Jonctiunile
de comunicare
Permit trecerea unor molecule mici dintr-o celula in
alta si sunt de de doua tipuri: jonctiunile permeabile si
sinapsele.
Jonctiunile permeabile, de tip gap, nexus sau macula comunicans sunt realizate prin structuri proteice, denumite conexoni,
ce strabat plasmalema, proeminand de o parte si de alta a
bistraturilor lipidice ale membranelor adiacente. Sunt raspandite in diferite tipuri de
tesuturi, reprezentand principala cale de comunicare intercelulara.
Permit schimburi rapide de molecule, facand posibila o cooperare
metabolica eficienta.Spatiul intercelular (spatium intercellulare) este foarte ingust (2-4nm). In momentul
realizarii jonctiunii permeabile, conexonii din cele doua
plasmaleme se aseaza cap la cap, formand canale directe de comunicare
intre citoplasmele celor doua celule, fara deschideri in
spatiul intercelular.(fig.3.18).
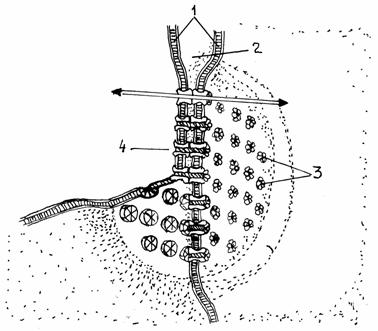
Fig.3.17Schema
jonctiunii permeabile (gap)
Plasmalemele
adiacente;
2-Spatiu intercelular;
3-Conexoni;
4-Canale intercelulare.
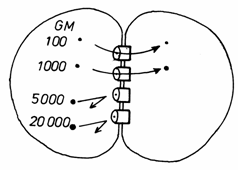
Fig.3.18.Diagrama
schimburilor moleculare prin jonctiunile gap.
Un conexon are o forma
de prisma hexagonala cu diametrul de 7-8 nm si este alcatuit din 6 subunitati proteice
(oligomeri). Subunitatile delimiteaza un canal hidrofil cu
diametrul reglabil. Ionii de calciu intracelular (Ca2+)
modifica diametrul canalului, controland astfel comunicarea intercelulara.
Canalul hidrofil al jonctiunii gap permite trecerea
dintr-o celula in alta a ionilor si a unor molecule cu greutate sub
1.000-1500 daltoni, precum: glucide, aminoacizi, nucleotide, hormoni,
vitamine,etc. Nu pot sa treaca macromoleculele (proteine, acizi
nucleici, etc.), dar sunt de 10.000 ori mai permeabile pentru ionii metalici
decat restul suprafetei membranei.
In culturile de celule, jonctiunile de tip gap se
organizeaza foarte rapid, din proteinele existente in plasmalema, care se grupeaza in conexoni. Ele
permit schimbul unor molecule de marimi medii, ca nucleotizii.
Jonctiunile gap sunt necesare mai ale in
situatiile in care celulele trebuie sa actioneze simultan, in
grup, cum este cazul celulelor
embrionare, pe cale de diferentiere sau intre celulele musculare (cardiace
si netede), nervoase si endocrine.
3.3.4.Complexele
jonctionale
Cuprind jonctiuni stranse spre frontul luminal
si desmozomi spre frontul latero-bazal. Sunt intalnite, mai ales, intre
celulele din epiteliile intestinale si renale. In complexele
jonctionale sunt prezente toate tipurile de legaturi intercelulare
necesare pentru indeplinirea functiilor specifice tesuturilor respective.(fig.3.19)
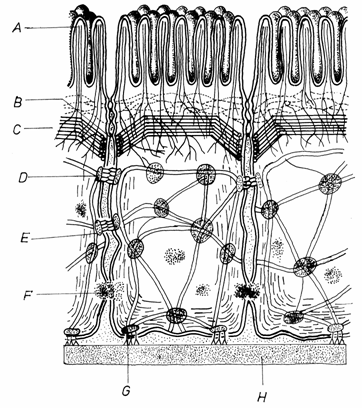
Fig.3.19. Complexe jonctionale in epiteliul intestinului subtire.
A-Micvrovili;
B-Jonctiuni impermeabile; C-Desmozomi in banda; D-Desmozomi in pata; E-Filamente
din citoschelet; F-Jonctiuni permeabile;
G-Hemidesmozomi; H-Mebrana bazala.
3.5. Receptorii din membrane
Receptorii din membrane sunt dispozitive moleculare
proteice intramembranare, cu ajutorul carora se intercepteaza
semnalele, ce sosesc pe cale nervoasa sau umorala.
Toate substantele care se leaga de receptori
si moduleaza (modifica) functia celulara se numesc liganzi.
Liganzii sunt substante, produse de celule specializate, care
actioneaza in mod specific asupra unui grup de celule
'tinta', producand-le modificari ale activitatii . Ei sunt
denumiti si mesageri extracelulari sau mesageri de ordinul I, cei mai
cunoscuti fiind hormonii si neurotransmitatorii.Liganzii
actioneaza in cantitate foarte mica (10-8 M), iar
receptorii ii leaga cu o constanta
de afinitate mare (K=108 litri/mol).Exista doua mari categorii de receptori in membrane: receptori pentru
substante endogene si receptori pentru substante exogene.
3.5.1. Receptorii
pentru substante endogene
Se cunosc mai multe categorii de molecule semnal (de liganzi) de
natura endogena (produse de organism): 1-mediatori chimici locali;
2-hormoni; 3-neurotransmitatori; 4-substante imunogene.
Mediatorii chimici locali sunt secretati de unele tipuri de
celule si actioneaza numai asupra celulelor din imediata lor
vecinatate. Ei sunt rapid captati si distrusi.(Fig.3.20).
Hormonii sunt substante
endogene produse de celulele glandelor endocrine. Pe cale sanguina, ajung
la celulele tinta din diferite organe, actionand la
distante mari de locul de producere.
Neurotransmitatorii
sunt substante endogene produse de neuroni si eliberate la
nivelul sinapselor chimice. Ei actioneaza numai asupra neuronilor
agajati in sinapsa.
Hormonii si
neurotransmitatorii sunt considerati mesageri de ordinul I.
Hormonii au o compozitie chimica variata, putand fi
substante steroide (hormonii sexuali) sau polipeptide ( ca de expl.
hormonii pancreatici si tiroidieni). Hormonii steroizi patrund in
celula, traversand bistratul lipidic, pe cand cei polipeptidici se
cupleaza cu receptorii specifici din membrana, actionand de la
acest nivel.
Substantele
imunogene sunt reprezentate de : -antigene endogene; - anticorpi; -
componentele complementului.
Receptorii pentru
neurotransmitatori sunt situati in membrana
postsinaptica a celulelor nervoase, in membrana celulelor musculare sau a altor celule efectoare. Ei
receptioneaza acetilcolina, noradrenalina, dopamina, serotonina,
adrenalina, histamina, acidul gama-aminobutiric, acidul aspartic,
encefalina,etc.
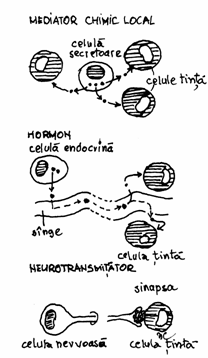
Fig.3.20.
Molecule semnal endogene.
Receptorii pentru
hormoni au o localizare diferita in celula, in functie de
tipul hormonului, hidrofil sau hidrofob. Hormonii hidrofili (insulina,
glucagonul, adrenalina, hormonii hipofizari, parathormonul,etc.) se
ataseaza de receptorii din
membrana periferica, transmitand celulei informatia necesara
pentru a-si modula activitatea. Dintre receptorii hidrofili, cei mai bine
studiati sunt receptorii pentru insulina din membrana hepatocitelor
si adipocitelor. Ei au o masa moleculara de 300.000 daltoni, iar
in membrana unei celule exista circa 10.000 receptori pentru
insulina. Hormonii hidrofobi (steroizi si tiroidieni)
traverseaza membrana celulei tinta si se leaga de
receptorii membranelor intracelulare.
Receptorii pentru
substantele imunogene sunt de trei feluri: 1-pentru antigene endogene;
2- pentru anticorpi; 3- pentru complement.
Receptorii pentru antigene endogene sunt mai
numerosi pe membrana limfocitelor T, sunt codificati genetic si
au capacitatea sa recunoasca structurile moleculare 'self'
(proprii organismului respectiv).Ei pot fi prezenti si in membrana
unor celule neimunogene (expl. in membrana enterocitelor.)
Receptorii pentru anticorpi sunt receptori Fc, pentru fractiunea
cristalizabila a imunoglobulinei G. Ei sunt prezenti in membrana
celulelor din sistemul fagocitelor mononucleare. In membrana bazofilelor si mastocitelor se
gasesc receptori pentru imunoglobulina E.
Receptorii pentru complement leaga cea de a treia
componenta a complementului de suprafata celulei, fiind
caracteristici celulelor din sistemul fagocitelor mononucleare.
3.5.2.Receptorii
pentru substante exogene
Sunt receptori pentru:
- virusuri, capabili
sa recunoasca adenovirusuri, mixovirusuri,etc.;
- antigene nonself (straine
de organism); sunt prezenti pe suprafata limfocitelor B si au o
structura moleculara asemanatoare
imunoglobulinelor.Acesti receptori determina transformarea
blastica a limfocitelor B, dupa contactul cu antigenul.
- toxine
microbiene. Sunt prezenti in membrana celulelor imunocompetente.
- lectine.
Lectinele sunt substante proteice sau glicoproteice, extrase din
tesuturi vegetale sau animale, capabile sa formeze punti intre
glicoproteinele membranare.
3.5.3.Mecanismul
de actiune al receptorilor.
Receptorii din membranele de suprafata
sunt activati in majoritatea cazurilor de molecule semnal (liganzi)
hidrofile.
Moleculele semnal hidrofile nu pot strabate membrana
celulara decat dupa ce se leaga de receptorii specifici. Sub
influenta diferitilor liganzi se produc modificari metabolice in
celulele tinta. Atasarea liganzilor de receptori se realizeaza
prin legaturi hidrofobe si legaturi de hidrogen ( ca de exemplu
legarea insulinei de receptori ). Legaturile sunt labile si strict
dependente de concentratia liganzilor din mediul extracelular.
Efectele actiunii complexului ligand-receptor asupra
celulelor constau in: modificari
structurale ale membranei celulare si modificari functionale ale membranelor.
Modificarile
structurale ale membranei celulare se manifesta prin redistribuirea receptorilor, raspanditi
la intamplare inainte de interactiunea ligand receptor. Dupa
contactul ligand-receptor, receptorii se grupeaza la suprafata
membranei in zone delimitate sau 'plaje'.(Fig.3.21)
Acest lucru se poate realiza datorita
fluiditatii bistratului lipidic, incat mai multe molecule de ligand
pot actiona asupra mai multor receptori invecinati. Astfel in
limfocitele B, lectinele pot induce formarea unor agregate receptor-ligand de
dimensiuni mari, denumite cupole, unde se declanseaza un proces de
endocitoza.
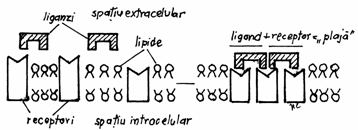
Fig. 3.21. Redistribuirea receptorilor din membrana
celulara
Modificarile
functionale ale membranelor constau in:
1- Modificari de permeabilitate a membranei,
caracteristice pentru liganzi de tipul neurotransmitatorilor. In
acest sens, acetilcolina se leaga de receptorii din membranele postsinaptice
ale fibrelor musculare, determinand cresterea permeabilitatii
membranei pentru ionii din mediul extracelular.
2- Inducerea de endocitoza, care se produce cand
ligandul este o substanta endogena, vehiculata pe cale
umorala.
3- Patrunderea din mediul extracelular a unor ioni
cu functie de mesageri de ordinul II. Astfel, ionii de calciu patrund
in celula dupa ce neurotransmitatorii se leaga de
receptorii din membranele post sinaptice ale jonctiunii neuromusculare.
4- Activarea unor enzime din membrana celulara. Se
produce cand liganzii sunt hormoni hidrofili. Asfel se activeaza adenilat ciclaza ce va cataliza sinteza,
pe fata interna a membranei, a adenozin 3 monofosfatului
ciclic (AMP-c) sau va determina fosforilarea proteinelor celulare. Activarea adenilat ciclazei se realizeaza cu
ajutorul unor proteine reglatoare.(Fig.3.22)
Dupa activarea adenilat ciclazei, in citosol
creste cantitatea de AMP-c (considerat mesager de ordinul II) ,
amplificandu-se semnalul hormonal. Totodata se activeaza si o
proteinkinaza care controleaza fosforilarea mai multor molecule
proteice, stimuland procesele metabolice din celula. Sinteza de AMP-c
poate fi blocata de unele prostaglandine care actioneaza prin
inhibarea actiunii adenilat ciclazei.
In unele maladii este implicata, in mod direct
disfunctia receptorilor din
membrana. Astfel, in 'miasthenia gravis' este inactivat
receptorul pentru acetilcolina de la nivelul placii neuromusculare,
iar in hipercolesterolemia genetica sunt absenti sau
nefunctionali receptorii pentru colesterol.
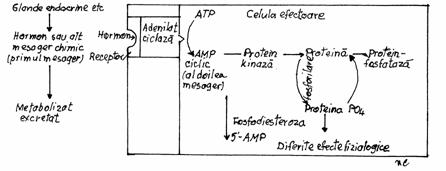
Fig.3.22.
Activarea unor enzime din membrana celulara
Exista afectiuni ale receptorilor care constau
in blocarea functiei autoimune prin autoanticorpi antireceptori. Asfel,
autoanticorpii pentru receptorii de insulina pot actiona ca
insulinomimetici (activand receptorii) sau ca blocanti ai acestora (in diabetul insulinorezistent).
Autoanticorpii care stimuleaza receptorii dopaminei pot constitui cauze
ale scizofreniei, iar imunoautoanticorpii beta adrenergici sunt implicati
in afectiunile asmatice. In procesul de imbatranire si in unele
boli neuropsihice (la om) s-a observat o modificare a densitatii
receptorilor.
3.6.Schimburile
prin membrana
Prin membrana celulara se efectueaza
schimbul de substante, fie prin traversarea diferitelor structuri ale
acesteia, sau pe calea transportului in masa, cu ajutorul veziculelor,
furnizate de membrana celulara.
bistratul
lipidic functioneaza ca o bariera de difuziune pentru apa
si alte molecule hidrofile, in timp ce moleculele liposolubile, oxigenul
si bioxidul de carbon il trec cu usurinta.
3.6.1.Transportul
transmembranar
Realizeaza schimburile
de molecule mici si ioni (de unde
si denumirea de microtransport) intre celula si mediul
extracelular. Indeplineste urmatoarele functii:
Preia din mediul
extracelular o serie de combustibili metabolici necesari mentinerii vietii celulei si
activitatilor ei metabolice;
Elimina in
mediul extracelular compusi fiziologici necesari organismului
(substante secretate), precum si substante toxice toxice ,
rezultate din activitatea celulei;
Regleaza
volumul celulelor;
Asigura
mentinerea ph-ului intracelular
si compozitia ionica la valori cat mai favorabile
desfasurarii activitatii metabolice celularte;
Produce
diferente de concentratii (gradiente ionice), care permit
desfasurarea unor activitati biologice complexe, cum sant
excitabilitatea nervului si a muschiului.
In functie de mecanismele care intervin in miscarea moleculelor prin membrana celulara se deosebesc
doua forme de transport : a-transportul activ si b- transportul
pasiv. (Fig.3.23).
3.6.11.Transportul
pasiv
Realizeaza trecerea moleculelor mici in sensul gradientului de concentratie, iar a
ionilor in sensul gradientului electrochimic. Se face fara consum de
energie, fiind independent de
metabolismul celular.
Exista doua modalitati de realizare a
transportului pasiv prin: difuziune simpla si prin difuziune facilitata.
Transportul pasiv
prin difuziune simpla. Se face lent, intrucat restructurarea
permanenta a membranelor biologice este o dificultate pentru
difuziune.Este supus legilor difuziunii si osmozei, fiind dependent numai
de forte fizice.(Fig.3.23)
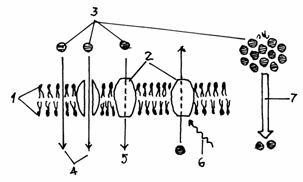
Fig.3.23. Transportul transmembranar.
1-Bistrat lipidic;2-Proteine
transmembranare;
3-Molecule transportate;4-Difuziune
simpla;5-Difuziune facilitata; 6-Transport activ;
7-Sensul
gradientului electrochimic.
Moleculele mici
liposolubile difuzeaza prin bistratul lipidic, iar cele hidrosolubile trec
prin orificii mici canale sau pori
hidrofili situati in bistrat, delimitati de proteine transmembranare.
Canalele transmemebranare au un diametru de 1 nm, fiind specializate si
selective. Vasopresina stimuleaza transportul prin canale si pori marindu-le
diametrul pana la 2-3 nm.
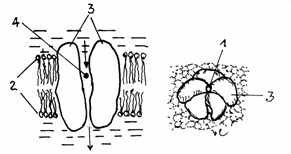
Fig.3.24.
Transportul prin canale si pori.
1-Por; 2-Bistrat lipidic; 3-Proteine transmembranare;
4-Ion.
Transportul prin
difuziune faciliata. Este mult mai rapid (de cca.100.000 ori)
decat cel prin difuziune simpla, permitand trecerea prin
membrana a unor substante greu solubile in lipide si cu o
masa moleculara relativ mare. Se intalneste frecvent la
hepatocite,unde, in acest mod,
se transporta
glucoza,aminoacizi,etc.(Fig.3.24.)
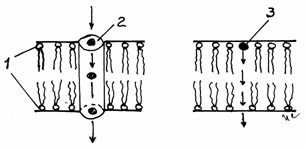
Fig.3.25.Tranportori
de tip canal si caraus.
1-Bistrat lipidic; 2-Canal;
3- Caraus sau tranportor mobil.
Se realizeaza cu
ajutorul unor molecule proteice din compozitia membranei cu rol de
carausi. Astfel, pe membrana hepatocitului exista 800.000
de carausi, iar fiecare caraus transporta
100 molecule de glucoza pe secunda.
Transportul prin difuziune facilitata se
efectueaza in sensul gradientului de concentratie si duce la
echilibrarea concentratiilor pe ambele fete ale membranei, insa viteza de transport este
direct proportionala cu concentratia lor. In raport cu
numarul speciilor transportate exista transport uniport, simport
si antiport.
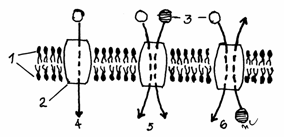
Fig.3.26.
Tipuri de transport, in functie de sens si numarul speciilor
transportate
1-Bistrat lipidic;
2-Proteina transportor; 3-Molecule transportate; 4-Uniport:
5-Sinport;6-Antiport.
3.6.1.2.Transportul
activ
Se realizeaza impotriva gradientelor de
concentratie sau a gradientelor electrochimice, producand o crestere
a concentratiei pe o fata a membranei. Solicita
cheltuiala de energie, care este furnizata de metabolismul celular,
fiind metabolic dependent. Unele celule inalt diferentiate (fibre musculare, neuroni) isi procura
energia necesara transportului pe cale aeroba, prin producere de ATP.
Hematiile isi produc moleculele de
ATP necesare pentru transport prin glicoliza.
Exista mai multe tipuri de transport activ, in
functie de modul in care este folosita energia:
1-Tranportul activ prin pompe ionice, in care proteinele
ce efectuiaza transportul folosesc direct energia din ATP, avand
proprietati ATP-azice ( de descompunere a ATP-ului).
2-Trasportul cuplat (sau cotransportul), in care se
foloseste energia gradientelor ionice, substantele de
transportat (glucide, aminoacizi) fiind
cuplate cu unii ioni (Na
3-Translocarea de grup, intalnita la unele bacterii,
care folosesc ca sursa de energie fosfoenolpiruvatul.
Transportul prin
pompe ionice.
Pompele ionice sunt
protein-enzime din compozitia plasmalemei care transporta ionii
intr-o directie termodinamica nefavorabila, impotriva
gradientului electrochimic sau de concentratie.
In organism se cunosc pompe transportoare pentru ioni sau
cationi,existand pompe pentru un singur ion (Ca2+sau Mg2+)
sau pentru doi ioni (pompa de Na+si K+).(Fig.3.26)
Pompa de Na+
- K+ este ce a mai bine cunoscuta.
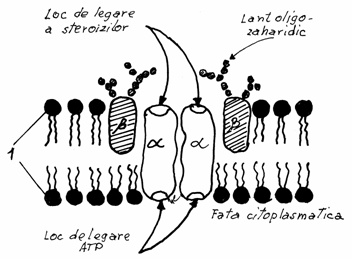
Fig.3.27.
Organizarea moleculara a pompei ionice de Na+ - K+
1-Plasmalema;
a -Proteine transmembranare
In conditii normale, ionul de potasiu (K+)
se gaseste intr-o concentratie mai mare (de cca. 15
ori)intracelular, in timp ce ionul de sodiu (Na+) are o
concentratie mai mare extracelular. Gradientele ionice (diferentele de concentratii) sunt
mentinute printr-un sistem special de transport, denumit pompa de Na+-K+.
Pompa de Na+-K+ a fost
studiata pe membranele hematiilor, unde s-a constatat o deplasare
concomitenta a Na+ extracelular (spre o concentratie mai
mare), iar a K+ intracelular (tot spre o concentratie mai
mare), contrar gradientelor de concentratie. Energia necesara acestui
transport se obtine prin hidroliza
ATP-ului, care este realizata de catre o enzima din
plasmalema, denumita Na+-K+-ATP-aza,
deoarece actioneaza numai in prezenta ionilor de Na+,K+
si a unei concentratii favorabile de Mg2+. Aceasta enzima este o
glicoproteina mare, un tetramer cu o greutate moleculara de 270.000 daltoni.
Pompa de Ca2+
sau Ca2+-ATP-aza
asigura transportul ionilor de Ca2+ prin sistemul de membrane
ale reticulului endoplasmic din fibrele musculare.
Exista si pompe pentru anioni, precum pompa de
clor (Cl-) din celulele parietale ale glandelor gastrice sau pompa
de iod (I-) din tiroida.
Transportul cuplat (sau
mecanismul de pompare moleculara) utilizeaza, pentru transportul unor molecule (glucoza,
aminoacizi), energia care rezulta din miscarea Na+
impotiva gradientului de concentratie si al gradientului electric.
Aceasta energie poate fi folosita pentru introducerea in celula
a unor substante nutritive. (Fig.3.28).
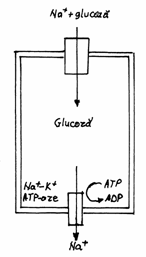
Fig.3.28.Transportul cuplat prin pompare moleculara.
Glucoza este pompata din mediul extracelular in
celule (enterocite sau nefrocite) odata cu ionii de Na + cu
ajutorul unei unitati proteice de transport. Dupa
patrunderea in celula, glucoza se decupleaza de ionii de Na+ , care sunt pompati in
afara celulei printr-o Na+-K+-ATP-aza.
3.6.2.
Transportul in masa
Este procesul prin care
celulele preiau din mediul extracelular particule de natura diferita,
prin formarea de vezicule pe seama membranei celulare. In functie de
directia in care se deplaseaza veziculele deosebim trei tipuri de
transport in masa: 1-exocitoza; 2-endocitoza; si 3-transcitoza.
3.6.2.1. Exocitoza
Exocitoza este procesul prin
care celulele isi descarca in mediul extracelular produsele secretate
sau alte materiale pemtru export. Produsele pentru export se colecteaza in vezicule
intracitoplasmatice (vesicula
cytoplasmatica, care se deplaseaza dinspre complexul Golgi spre
membrana celulara, fie in mod spontan, fie dupa stimularea celulei de
catre unii factori secretagogi (hormoni, neurotransmitatori,
factori de eliberare). Deschiderea veziculelor si eliminarea
continutului in spatiul extracelular se datoreste contractiei
microfilamentelor, in prezenta moleculelor de ATP , de AMP-c sia ionilor de Ca2+.In
celulele exocrine, exocitoza este limitata la domeniul apical (luminal) al
plasmalemei.
Fig.3.29.Transportul
prin vezicule (in masa).
A-Endocitoza; B-Exocitoza;
C-Transcitoza.
1-Plasmalema;
2-Fagozom; 3-Lizozom primar; 4-Lizozom secundar;5-Vezicule de secretie; 6-Vezicule de
transport.
Exocitoza este prezenta si la nivelul
sinapselor, unde veziculele ce contin mediatori chimici
(acetilcolina, noradrenalina) fuzioneaza cu membrana
presinaptica. Mediatorii sunt
eliberati in spatiul sinaptic, dupa care se leaga de
receptorii specifici din membrana postsinaptica, producand depolarizarea
ei si transmiterea influxului nervos.
3.6.2.2. Endocitoza
Endocitoza este procesul
prin care celulele preiau din mediul extracelular molecule mari ( cu greutate
moleculara mai mare de 10.000 daltoni), prin intermediul unor vezicule,
care se formeaza din membrana celulara. In acest fel se poate realiza
nutritia celulei, purificarea mediului extracelular si unele procese
de aparare imunitara.
Dupa natura materialului endocitat si dupa
modaliatea de ingestie se disting
doua tipuri de endocitoza: 1-fagocitoza
(de la grecescul phagein= a
manca) sau endocitoza de molecule si
particule solide,microorganisme (bacteriene sau virale), macromolecule
alterate, detritus celular, celule intregi, etc. neinsotite de fluid; 2-
pinocitoza ( de la grecescul pinein= a bea) sau endocitoza de fluid, ce
contine molecule sau particule.
3.6.2.2.1.Fagocitoza
In conditii normale
si patologice, pot participa la fagocitoza un mare numar de tipuri celulare,
denumite generic fagocite. In functie de marimea particulelor pe care sunt
capabile sa le inglobeze, se disting microfage,
care inglobeaza numai particule mici (exemplu neutrofilele) si macrofage, care inglobeaza
particule mari, existand macrofage mobile ( monocitele circulante) sau
macrofage fixe (histiocitele din tesutul conjunctiv, celulele Kupffer din
ficat, celulele capilarelor sinusoide din splina, unele celule din
maduva osoasa hematogena, din limfonoduri, din pulmon,etc.).
Fagocitele prezinta pe suprafata membranei
regiuni specializate purtatoare de receptori, cu ajutorul carora
recunosc ceea ce este 'self' (macromoleculele proprii organismului)
de ceea ce este ''non self', adica macromoleculele
straine de organism, denumite generic antigen. Dintre macromoleculele self,
receptorii pentru fagocitoza rercunosc ceea ce este sanatos de
ceea ce este alterat ( self alterat: celule degenerate, celule maligne, celule
imbatranite, resturi tisulare). In unele situatii patologice,
receptorii celulelor ce intervin in fagocitoza pierd capacitatea de a
diferentia self-ul de non-self , actionand, asfel, impotriva proteinelor
din organismul propriu si determinand aparitia reactiilor
autoimune, care pot genera o serie de entitati patologice.
Recunoasterea antigenilor in organism este
inlesnita de prezenta in mediul extracelular a unor proteine,
denumite opsonine. Opsoninele sunt proteine extracelulare care mediaza
si faciliteaza legatura intre receptorii membranei fagocitelor si antigenele ce vor fi
fagocitate, formand complexe antigen-opsonine.
Se cunosc mai multe tipuri de receptori ai complexelor
antigen-opsonina, precum: - receptorii Fc, care leaga de suprafata membranei
antigenele opsonizate cu imunoglobuline G ( Ig G), dupa ce au recunoscut
fractiunea cristalizabila (Fc) a imunoglobulinei G.; - receptorii C3, care reconosc si fixeaza
antigenii opsonizati cu cel de al treilea component al complementului.
In membrana fagocitelor, in afara de receptoriii
pentru complexe opsonizate, s-au mai descris receptori nespecifici care
recunosc si fixeaza selful alterat, reprezentat de resturi celulare
sau celule maligne.
Fagocitoza se desfasoara in patru
etape:chemotaxia,opsonizarea, ingestia si digestia.
Chemotaxia este proprietatea unei celule mobile (in czul de fata a
neutrofilelor) de a se deplasa prin tesuturi catre particule
tinta, ca raspuns la diversi stimulli dupa ce
tinta a fost recunoscuta.
Semnalele chemotactice sunt reprezentate de: substante bacteriene
(proteine, lipide), proteine serice, componente ale complementului (15 proteine serice, care amplifica
raspusul imun), produse ale limfocitelor (limfokine), factori
eliberati de neutrofile.
Opsonizarea consta in
acoperirea bacteriilor sensibilizate de anticorpi pentru a fi fagocitate.
Principalele opsonine sunt anticorpii, in special imunoglobuline (Ig1,
Ig3) si componente
ale complementului (C3, C5).
Fractiunea cristalizabila (Fc) a imunoglobulinei G este
recunoscuta de receptorii Fc de pe suprafata neutrofilelor. (Fig.3.30)
Fig.3.30.
Opsonizarea si fagocitarea.
Ingestia microorganismelor are loc dupa ce
acestea au fost opsonizate. Complexul antigen-opsonine este recunoscut de receptorii
de la suprafata fagocitelor si legat de membrana celulara,
determinand activarea receptorilor. In urma activarii receptorilor, actina
din citoscheletul membranei,se contracta, determinand emiterea de pseudopode. Pseudopodele inconjoara strans
particula si in acest fel se realizeaza interactiuni receptor-
particula pe toata suprafata ei. Mecanismul de fixare a
particulei de membrana psedupodelor este comparat cu inchiderea unui fermoar.
Extremitatile pseudopodelor inconjoara particula si
formeaza o vezicula, denumita fagozom.(Fig.3.31.)
In citoplasma,
fagozomul se uneste cu un lizozom , formandu-se un fagolizozom, in care
hidrolazele acide vor actiona asupra particulei straine, producand
digestia acesteia. In cazul
neutrofilellor, contopirea granulelor azurofile primare (a lizozomilor) cu
fagozomul si eliberarea continutului lor in fagolizozom (sau vezicula
digestiva) se manifesta prin
disparitia granulelor din citoplasma,fenomen cunoscut ca
degranularea neutrofilelor. 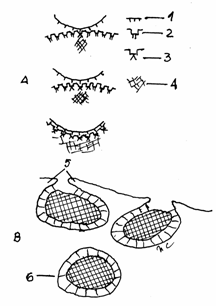
Fig.3.31.Mecanisme le fagocitozei.
A-Mecanisme moleculare;
B-Realizarea fagozomilor.
1- ImunoglobulinaG;
2-Receptor inactiv;
3-Receptor activat;
4-Proteina
contractila din citoschelet;
5-Pseudopode;
6-Fagozom.
Digestia intracelulara a
particulelor fagocitate are loc in lizozomi si se produce prin: -sisteme
dependente de oxigen, mediate de enzime (mieloperoxidaza,
superoxidismutaza); si - prin sisteme independente de oxigen ,ce
presupun actiunea lizozimului, a lactoferinei,etc..
3.6.2.2.2.Pinocitoza
sau endocitoza particulelor in faza fluida.
Pinocitoza reprezinta
procesul de transport in masa a unei cantitati variabile de
fluid tisular, impreuna cu particulele pe care le contine, prin
intermediul unor vezicule (vezicula
pinocitocica), denumite pinozomi.
Se
intalneste la toate tipurile de
celule, aparand ca o modaliatate importanta prin care celulele capteaza din lichidul
intercelular substantele necesare metabolismului lor.
Se deosebesc doua forme de pinocitoza in
functie de mecanismele care intervin in procesul de preluare a
sustantelor: - pinocitoza fara receptori si - pinocitoza
mediata de receptori.
Pinocitoza
fara receptori este
endocitoza cea mai frecvent intalnita la acele celule din organism, care
folosesc pentru transport suprafete mari nespecializate din membrana
plasmatica.. Preluarea substantelor cu ajutorul veziculelor se face
fara o legare prealabila
a lor de membrana celulara prin
receptori.
Pinocitoza se desfasoara in mai multe
etape:
1-Contactul particulelor din fluidul tisular produce
activarea unor puncte de pe
suprafata celulei , denumite situsuri
anionice. Acest proces induce agregarea microfilamentelor din citoscheletul
membranei, producand cresterea flexibilitatii membranei
celulare.
2-Membrana celulara se invagineaza formand
cripte sau canale intracelulare
(invaginatio cellularis), in care intra lichid extracelular,
impreuna cu particulele prezente in el.
3-Membranele din vecinatatea extremitatii
profunde a canalului se alipesc,fuzioneaza si apar pinozomii (vesicula pinocytotica), vezicule ce se desprind de canal si sunt antrenate de curentii intracitoplasmatic.
4-Pinozomii fuzioneaza cu lizozomii, formand pinolizozomii, in care enzimele
lizozomale produc digestia materialelor preluate din exteriorul celulei.
In
functie de marimea particulelor inglobate prin pinocitoza se
distinge o micropinocitoza si o-macropinocitoza.
Macropinocitoza
este modalitatea de transport a unor particule mari vizibile la microscopul
optic. Poate fi studiata prin injectarea "in vivo" de coloranti vitali, care se fixeaza de proteinele
serice. Complexele formate din proteine serice si colorantii vitali
coloidali sunt introduse in citoplasma impreuna cu mici
cantitati de lichid
extracelular, fiind utilizate ca
indicator de recunoastere a celulelor care manifesta proprietate de
pinocitoza la nivelul organelor si tesuturilor.
Micropinocitoza reprezinta modalitatea de transport a
moleculelor mici cu ajutorul veziculelor , vizibile numai la microscopul
electronic.
Urmarindu-se soarta veziculelor pinocitate, s-a
observat ca acestea pot: 1- ramane in citoplasma, unde
fuzioneaza cu lizozomii, in care sunt digerate, iar dupa ce
si-au descarcat continutul, membranele lizozomale sunt
reintegrate in membrana celulara; 2- traversa citoplasma fara a
fuziona cu lizozomii, descarcandu-si continutul pe cealalta
fata a citoplasmei.
Pinocitoza mediata de receptori.
Se mai numeste si
pinocitoza absorbtiva, selectiva sau concentrativa. Este
modalitatea de preluare din mediul extracelular a unor materiale diferite, prin
folosirea unor zone specializate ale membranei celulare, prevazute cu
receptori.
Receptorii recunosc si leaga de membrana
particule diferite. Fiind mobili receptorii se grupeaza la nivelul unor
zone specializate ale membranei celulare, care se invagineaza formand caveolele (vesicula superficialis s.
caveola) acoperite, deoarece sunt acoperite pe versantul citoplasmatic de o
retea de microfilamente, formate dintr-o proteina, denumita clatrina. Reteua de
clatrina stabilizeaza pozitia receptorilor si mentin
caveolele in comunicare cu mediul extracelular.(Fig.3.32.)
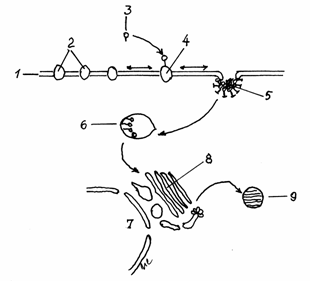
Fig.3.32. Pinocitoza mediata de receptori.
1-Plasmalema;
2-Receptori mobili;3-Ligand; 4-Complex ligand receptor;
5-Caveola
acoperita cu clatrina; 6-Receptozom; 7-Nucleu; 8-Complex Golgi; 9-Lizozom.
Caveolele acoperite care fixeaza complexele
particula-receptor (sau ligand-receptor) sunt inglobate rapid
impreuna cu o cantitate mica de fluid extracelular sub forma de
vezicule care pot sa-si pastreze mantaua de clatrina pe
fata lor externa (vezicule acoperite) sau microfilamentele de
clatrina se desprind la nivelul membranei, in acest caz aparand
vezicule netede sau receptozomi.
Particulele inglobate prin pinocitoza mediata de
receptori pot fi degradate in lizozomi sau in unele cazuri, receptozomii
traverseaza citoplasma fara sa interactioneze cu
lizozomii, fiind eliminate pe cealalta fata a celulelor.
Spre deosebire de pinocitoza fara receptori, in
care concentratia substantelor in veziculele endocitate este aceeasi
ca in lichidul extracelular, in veziculele acoperite , ce apar in pinocitoza
mediata de receptori, concentratia substanltelor este mult mai
mare decat in lichidul extracelular, datorita capacitatii
receptorilor de a lega selectiv un numar mare de particule.
Implicatii
patologice. Pinocitoza mediata de receptori este o modalitate
folosita pentru preluarea din lichidul extracelular si din sange a
colesterolului, o lipoproteina cu densitate mica. In situatii
normale, celulele prevazute cu receptori pentru proteine cu densitate
mica preiau colesterolul din mediul extracelular. Lipsa acestor receptori,
intalnita in unele deficiente genetice, face imposibila
preluarea moleculelor de colesterol din sange, determinand o crestere a
concentratiei acestuia (hipercolesterolemie) si aparitia de
placi aterosclerotice in peretele vascular.
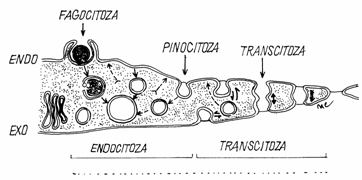
Fig.3.33.
Schematizarea modalitatilor de transport prin celula
endoteliala.
(dupa SIMIONESCU-1981)
3.6.2.3.Transcitoza
Transcitoza sau citopemsis reprezinta o forma
aparte a transportului prin vezicule,
prin care se realizeaza transportul macromoleculelor prin celule
endoteliului capilar. A fost studiata de Palade (1953), care a observat,
cu ajutorul microscopului electronic, vezicule ce traverseaza celulele
endoteliale,ralizand schimburi intre plasma si lichidul
interstitial.
Transportul veziculelor pinocitate se poate realiza prin
doua modalitati: 1-transcitoza distributiva, in care
trecerea se face sub forma unor siruri de vezicule intracitoplasmatice, ce
se intind de la fata luminala
la fata bazala, fara a ajunge in contact cu lizozomii;
si 2-transcitoza conectiva, in care transportul se face prin canale
ce se formeaza fie prin fiziunea veziculelor, fie prin invaginarea
plasmalemelor.

