COMPOZITIA
BIOCHIMICA A MEMBRANELOR BIOLOGICE
Pentru a intelege functiile membranelor
biologice este necesar a le cunoaste structura moleculara.
Componentele moleculare ale oricarei membrane biologice sunt proteinele, lipidele polare si glucidele
in alcatuirea glicoproteinelor si glicolipidelor.
Proportiile in care sunt reprezentate
diferitele componente biochimice variaza cu functia si
localizarea biomembranelor respective.
De exemplu, multi neuroni au extensiile
invelite de o teaca de mielina. Teaca de mielina care este
multistratificata este compusa in mare parte din lipide care induc
rolul de izolator electric. La nivelul membranelor mitocondriilor sau
cloroplastelor (echivalentul mitocondriilor la celulele vegetale), acolo unde
au loc numeroase reactii catalizate enzimatic, proteinele sunt majoritare.
Fiecare specie, fiecare tip de tesut sau de
celula precum si diferitele tipuri de organite poseda un set
caracteristic de lipide membranare. De exemplu, plasmalema hepatocitelor este
bogata in colesterol si nu contine nivele detectabile de
cardiolipina. In acelasi timp, membrana mitocondriala
interna a hepatocitelor prezinta o distributie inversata.
Este evident ca in celula exista mecanisme de control a
cantitatii de lipide sintetizate si a modului de distributie.
Compozitia proteica a membranelor
variaza mai mult decat compozitia lipidica, reflectand
specializarea functionala (1 functie - 1 specie proteica).
De exemplu, la nivelul celulelor cu bastonas din retina, o
portiune a celulei este specializata pentru receptia luminii. La
acest nivel, peste 90% din compozitia plasmalemei este reprezentata
de o glicoproteina fotoabsorbanta - rodopsina. Plasmalema
eritrocitului, care are o specializare mai redusa fata de celulele
cu bastonas, prezinta circa 20 de tipuri de proteine esentiale
si alte cateva sute de importanta mai mica. Multe din
aceste proteine sunt transportori pentru solviti prin membrana.
Unele proteine membranare sunt legate covalent de
retele complexe de carbohidrati. De exemplu, glicoforina, o
glicoproteina din membrana eritrocitului este compusa in
proportie de 60% din unitati oligozaharidice complexe
atasate covalent la reziduuri specifice de aminoacizi (cele mai utilizate
puncte de legatura le reprezinta reziduurile Ser, Thr si
Asn). Ca si exemplu antinomic prezentam rodopsina, glicoproteina
majoritara din membrana celulelor cu bastonas din retina care
prezinta un singur hexazaharid. Componentele glucidice de la
suprafata glicoproteinelor influenteaza plierea lanturilor
de aminoacizi precum si stabilitatea si destinatia
intracelulara; joaca de asemenea un rol esential in cuplarea
specifica a liganzilor la nivelul receptorilor glicoproteici de
suprafata. Membranele intracelulare (de exemplu, membranele mitocondriale,
contin rareori componente carbohidrate cuplate. Unele proteine sunt
cuplate covalent la unul sau mai multe lipide care joaca rolul de ancore
hidrofobe.
ARHITECTURA
SUPRAMOLECULARA A MEMBRANELOR BIOLOGICE
Membranele biologice:
sunt impermeabile pentru solvitii polari si
permeabile pentru solutiile non-polare;
au o grosime care variaza intre 5-8 nm (50-80
angstromi);
au un aspect trilamelar (trilaminar) pe sectiune,
prin vizualizare la microscopul electronic;
Studiile combinate de microscopie electronica
si de compozitie chimica, de studiu a permeabilitatii
si mobilitatii lipidelor si proteinelor a determinat
dezvoltarea modelului de mozaic fluid.
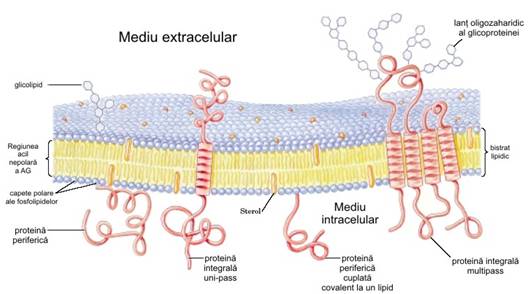
Conform acestui model, fosfolipidele si
sterolii formeaza un bistrat lipidic in care regiunile nepolare ale
moleculelor lipidice sunt dispuse in apozitie (fata in
fata) la nivelul miezului bistratului in timp ce capetele polare
privesc spre exterior. In acest bistrat, lipidele sunt inclavate la intervale
neregulate si mentinute prin interactiuni hidrofobe dintre
lipidele membranare si domeniile hidrofobe ale proteinelor. Unele proteine
protruzioneaza numai pe o singura parte a bistratului lipidic, altele
avand domenii de ambele parti ale acestuia. Orientarea proteinelor in
bistrat este asimetrica dand aspectul regionalizat al membranei. Domeniile
proteice expuse de o parte si de alta a bistratului difera, ceea ce
reflecta asimetria functionala. Modelul de mozaic fluid
reprezentat de lipidele si proteinele membranare are particularitatea
esentiala a libertatii de miscare laterala in
planul membranei.
LIPIDELE
MEMBRANARE
Lipidele reprezinta substante cu
semnificatie biologica particulara care sunt insolubile in
apa si solubile in solventi organici cum ar fi: acetona
(propanona), etanol, cloroform (triclorometan), dietileter (etoxietan) si
petrol usor (cu punct de fierbere la 40-60°C).
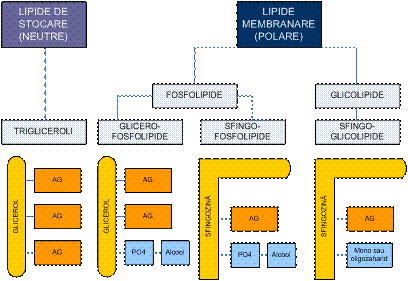
Din punct de vedere biochimic, lipidele se pot
clasifica in:
lipide de stocare - neutre
lipide membranare - polare
o
fosfolipide
glicero-fosfolipide
sfingo-fosfolipide
o
glicolipide - sfingo-glicolipide
o
colesterol
Clasificarea lipidelor (schema) - dupa Lehninger
Lipidele sunt molecule care confera membranei functia de
bariera. Hartley, in 1936, le-a descris ca si structuri amfifile sau
amfipatice. Aceasta inseamna ca lipidele sunt molecule asimetrice,
ele posedand un "cap" polar, hidrofil si o portiune alungita
"coada" hidrofoba.
Fosfolipidele
(sau fosfogliceridele)
Sunt cele mai abundente molecule lipidice. Ele
sunt compuse dintr-un cap polar si o coada non-polara. Ele
provin din esterificarea glicerolului (propan-1,2,3-triol) la nivelul grupelor
-OH. Aceste esterificari se fac la nivelul gruparilor OH din
pozitiile 1 si 2 cu acizi grasi iar la nivelul OH-ului din
pozitia 3 cu un derivat al acidului fosforic.

Natura substituentului X, fixat la nivelul
acidului fosforic precizeaza natura fosfogliceridului.
Cel mai simplu compus rezultat din esterificare
(in cazul in care X este hidrogenul -H) se numeste acid fosfatidic. Acesta
nu este un component al membranei, el aparand numai in cadrul sintezei
altor fosfogliceride. Cele mai importante fosfogliceride membranare sunt:
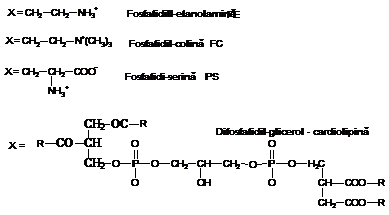
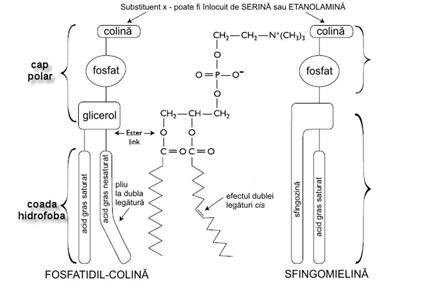
Sfingolipidele sunt lipide de o mare
diversitate, construite pe baza unui schelet constituit de o molecula de sfingozina care este un alcool
aminat cu lant lung de atomi de carbon.
Putem distinge 3 clase de sfingolipide:
Sfingofosfolipide (sfingomieline) Sfingomielinele joaca un rol
important la nivelul membranelor mielinice.
Glicosfingolipide neutre
Glicosfingolipidele
neutre
sau glicozil-ceramidele sunt formate
din atasarea la nivelul primului alcool al ceramidei, printr-o
legatura glicozidica, a glucozei, galactozei sau a unui alt
polizaharid (di, tri sau tetrahexoza). Putem astfel obtine o mare
diversitate de glicosfingolipide. Ele sunt denumite popular cerebrozide deoarece au fost descrise
prima oara la nivelul creierului (b-galactozil-ceramida);
ulterior ele au fost descrise si la nivelul altor tesuturi (b-galactozil-ceramida, in splina, la un
pacient cu maladie Gaucher).
Exemplificarea implicarii glicolipidelor
neutre in determinismul grupelor sangvine. Aceste glicolipide fac parte din
antigenele de suprafata care determina grupele sangvine la
suprafata hematiilor.
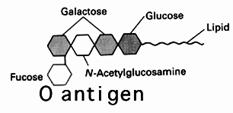
Grupa 0 poseda anticorpi
antiA si antiB iar antigenul specific este prezentat in figura de mai jos.
Cei cu grupa 0 pot primi sange numai de la grupa 0.
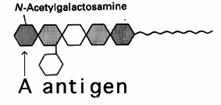
Grupa A poseda anticorpi
antiB iar antigenul specific este prezentat in figura de mai jos Acest antigen
este similar cu cel de la grupa 0 avand insa in plus o grupare
N-acetil-galactozamina. Cei cu grupa A pot primi sange de la grupa A
si 0.
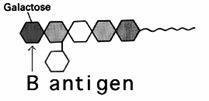
Grupa B poseda anticorpi
antiA iar antigenul specific este prezentat in figura de mai jos. Antigenul
este similar cu cel de la grupa A dar in loc de N-acetil-galactozamina
prezinta galactoza. Cei cu grupa B pot primi sange de la grupa B
si 0.
Grupa AB nu
poseda
anticorpi iar la suprafata hematiilor gasim ambele tipuri de
glicolipide de la grupele A si B. Cei cu grupa AB pot primi sange de la
toate grupele.
Glicosfingolipide acide
c. deriva din glicolipidele
neutre.
Cuprind 5 clase:
sialo-glico-sfingolipide (gangliozide - contin unul
sau mai multe reziduuri de acid sialic, denumit si acid N-acetil
neuraminic - NANA);
urono-glico-sfingolipide (contin unul sau mai multe
reziduuri de acid uronic);
sulfo-glico-sfingolipide (contin grupari
esterice carbohidrat-sulfat);
fosfo-glico-sfingolipide (contin grupari mono
sau diester-fosfat);
fosfono-glico-sfingolipide (contin grupari
2-aminoetil-hidroxifosforil)
Descoperite in celulele ganglionare (de unde
si denumirea de gangliozide),
ele reprezinta 6% din lipidele membranelor neuronale dar se gasesc
si in alte celule. Glicolipidele sunt acide, deci incarcate electric
datorita acidului sialic (care are o incarcare negativa). Rolul
lor nu se cunoaste precis (sunt presupusi receptori) dar exista
cateva exemple:
epiteliile de acoperire
contin o cantitate mare de gangliozide care protejeaza membrana de
actiunea degradativa a factorilor diversi, cum ar fi pH-ul,
enzimele litice.
prezenta gangliozidelor in membrane
poate influenta incarcarea electrica si captarea ionilor de
Ca2+.
toxina vibrionului holeric nu
poate penetra enterocitele decat in prezenta, la polul apical al acestor
celule, a unui gangliozid, denumit GM1. Patrunderea acestei
toxine in interiorul enterocitelor produce cresterea concentratiei de
AMPc care conduce la o secretie crescuta de Na si apa
catre lumenul intestinal, ceea ce produce diareea caracteristica.
Proprietatile lipidelor
membranare
Autoaranjarea spontana (autoorganizarea).
Aceasta proprietate se datoreaza caracterului amfifil (sau
amfipatic) descris; astfel, fosfolipidele si glicolipidele introduse in
apa formeaza un dublu-strat.
Din diverse experimente, cunoastem 2 tipuri de organizare a
bistraturilor lipidice:
bistraturi lipidice plane
(denumite si membrane negre);
bistraturi lipidice ce
formeaza vezicule - denumite liposomi.
o alta forma de
organizare spontana este reprezentata de micelii, de asemenea sferice
ca si liposomii, dar, spre deosebire de acestia, cozile lor hidrofobe
exclud apa din interiorul lor.
A. Bistraturile lipidice plane
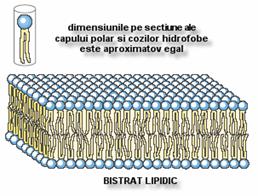
Pot fi obtinute prin amplasarea unei picaturi lipidice dizolvate
in solventi organici, la nivelul unui orificiu care separa 2
compartimente ce contin apa.
B. Liposomii
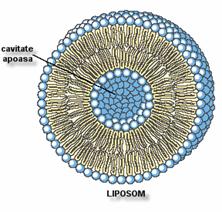
Fosfolipidele introduse in apa pot forma liposomi multilamelari, cu un
diametru de aproximativ 2 mm. Formarea lor poate fi
accelerata de agitarea mecanica a vasului de formare. Prin
tratamentul lor ultrasonic pot fi transformati in liposomi unilamelari.
Liposomii sunt folositi in experiente de reconstruire a membranelor
prin aditie de proteine. Astfel ei sunt utilizati in
terapeutica. Ei permit vehicularea (transportarea) unor medicamente pe care le inglobeaza, in torentul
sangvin de unde pot fi dirijati catre tesuturile
tinta.
C.
Miceliile
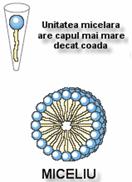
Miceliile reprezinta structuri sferice care contin cateva zeci
pana la cateva mii de molecule lipidice dispuse cu regiunile hidrofobe
agregate spre interior, excluzand moleculele de apa, si cu capetele
hidrofile spre exterior, in contact cu mediul apos.
Fluiditatea bistraturilor lipidice. Bistraturile lipidice
sunt structuri cvasifluide, interiorul bistratului fiind fluid. Aceasta
cvasi-fluiditate este rezultatul miscarilor efectuate in special de
fosfolipidele membranare. Cele mai importante miscari efectuate de
lipide sunt rotatia in jurul
propriei axe si miscarile
lanturilor laterale. Fluiditatea este proportionala cu
amplitudinea acestor miscari si are ca rezultat deplasarea
lipidelor si a proteinelor in planul membranei.
Miscarile
lipidelor sunt:
a. Intramoleculare - cand se
deplaseaza numai anumite portiuni ale moleculei lipidice.
b. De rotatie si de
translatie cand in miscare este angajata intreaga
molecula.
Miscarile
intramoleculare sunt putin accesibile si numai metodelor de investigatie ca
RMN (Rezonanta Magnetica Nucleara) sau RES (Rezonanta
Electromagnetica de Spin). Aceste miscari sunt reprezentate de
miscari ale grupurilor de atomi in jurul legaturilor simple
precum si de miscari segmentare la nivelul acizilor grasi
si modificari de conformatie la nivelul gruparilor polare.
Miscarile de
rotatie si de translatie sunt mai accesibile
investigatiilor.
Miscarea de rotatie este foarte rapida
(10-6-10-7 sec.). Moleculele se rasucesc in jurul
propriei axe si, din cauza flexibilitatii lanturilor
laterale, molecula lipidica va fi flectata la maxim la nivelul
miezului dublului strat lipidic si mai putin flectata la nivelul
capetelor polare.
Miscarile de
translatie sunt de doua tipuri:
w o miscare de
translatie laterala
w o migrare a lipidelor dintr-un
strat in altul.
Prin translatie laterala, lipidele isi schimba locurile
intre ele de circa 107 ori pe secunda, deci coeficientul de
difuziune (D) este in medie de 10-8 cm2/sec. Aceasta
inseamna ca o molecula lipidica poate difuza
('strabate') suprafata unei bacterii mari (aprox. 2 mm) intr-o secunda. Este permisa
comparatia cu un ocean lipidic in care moleculele componente au libertate
de miscare datorita interactiunilor Van der Waals.
Miscarile de lateralitate afecteaza si mobilitatea
proteinelor care, pentru regionalizare si pastrarea
polaritatii functionale necesita sisteme specifice de
ancorare prin generarea de clustere (agregate macromoleculare lipido-proteice
care pastreaza pozitiile relative ale proteinelor in membrana)
sau prin ancorarea la elementele citoscheletului cortical (cazul proteinelor
transmembranare). Un exemplu util este cel al glicoforinelor si pompelor
Na/bicarbonat care sunt cuplate si imobilizate de spectrina, o
proteina specifica citoscheletului eritrocitelor.
Asimetria dublului strat lipidic.
Un mare numar de membrane au o componenta lipidica
diferita pentru cele 2 monostraturi din structura. Originea
asimetriei chimice se gaseste la nivelul locului lor de sinteza,
adica in reticulul endoplasmic neted (un organit celular specializat),
unde se gasesc translocatori ai lipidelor care le insera si le
repartizeaza intr-un monostrat sau in altul (asa cum se va vedea in
capitolele referitoare la organitele celulare, reinnoirea membranelor celulare
depinde de reticulul endoplasmatic - atat rugos cat si neted - care
'inmugureste' trimitand vezicule spre membrana
celulara cu care fuzioneaza).

