SERIA
ERITROCITARA
ERITROPOIEZA
Eritropoieza reprezinta procesul
de diferentiere si maturare a precursorilor eritrocitari pana la
eritrocitele mature functionale, in paralel cu sinteza componentelor lor
specifice (hemoglobina). Dureaza 7 zile, iar in vitro,
2 saptamani.
Unitatea anatomica si
functionala a eritropoiezei este insula eritroblastica, vizibila in general pe biopsia de
maduva osoasa (MO); este formata din:
macrofag, care este situat central,
inconjurat de eritroblasti in diferite stadii de maturatie
(macrofagul trimite prelungiri care inconjoara fiecare eritroblast);
eritroblasti - cu o
dispozitie caracteristica: in interiorul insulei eritroblastice se
gasesc eritroblastii mai tineri, cei maturi fiind situati spre
exterior;
Intre eritroblasti si
macrofag exista anumite relatii - macrofagul fagociteaza nucleul
eritroblastului oxifil si o portiune din membrana acestuia, precum
si alte produse de degradare; eritroblastii preiau din macrofag
fierul (feritina) pentru sinteza hemoglobinei, precum si alte substante
nutritive. Practic, macrofagul este "doica" eritroblastilor.
Citodiabaza reprezinta trecerea
reticulocitelor din MO in sangele periferic, proces ce se face prin
miscari active, reticulocitul emitand un pseudopod care-l
impinge intre doua celule endoteliale. Aceasta prelungire devine din
ce in ce mai voluminoasa, pana cuprinde intreaga masa a celulei.
Reticulocitul intrat in torentul circulator isi recapata forma
obisnuita.
Maturatia seriei eritroide
consta in:
-
scaderea dimensiunilor celulei;
-
scaderea dimensiunii nucleului;
-
cresterea progresiva a cantitatii de citoplasma;
-
incarcarea progresiva a citoplasmei cu Hb ce duce la virarea culorii
din bazofila (albastra) in roz-portocalie, datorita componentei
oxifile (Hb).
Progenitorii celulari eritrocitari:
BFU-EB ("burst forming
unit-erythroid")
sunt cei mai tineri progenitori ai seriei eritrocitare, avand proprietatea de a
forma colonii eritroide de talie mare, cu caracter exploziv, independente de
eritropoietina si dependente de IL-3. Aceste celule prezinta
pe suprafata markeri HLA, antigene de grup sanguin slab reprezentate
si receptori pentru eritropoietina si transferina
CFU-E ("colony forming
unit-erythroid")
sunt progenitori mai diferentiati, dau nastere unor colonii
formate din 8-100 eritroblasti, dependenti de eritropoietina.
Coloniile se formeaza mai lent si au aceeasi markeri ca la
BFU-E, in plus antigenele de grup sanguin sunt mai bine reprezentate si au
un numar mai mare de receptori pentru eritropoietina.
Precursorii eritrocitari sunt celule identificabile morfologic.
Proeritroblastul este celula
cap de serie care se gaseste numai in MO, de talie mare (15-22 m),
forma rotunda sau ovalara si reprezinta aproximativ 1%
(0,5-1,5%) din elementele seriei rosii. Nucleul este mare, ocupand aproape
toata celula, forma rotunda, culoare albastru intens
datorita cantitatii crescute de ribonucleoproteine,
prezinta 1-2 nucleoli vizibili, are o zona clara
perinucleara; cromatina nucleara are aspect granitat. Citoplasma este
redusa cantitativ, bazofila, prezinta un halou perinuclear,
bogata in poliribosomi, mitocondrii in numar mare si activ
implicata in sintezele proteice (mai ales in sinteza Hb).
Eritroblastul bazofil are diametru de 12-18 m, nucleu
comparativ cu al proeritroblastului, dar ceva mai mic si cu cromatina
mai densa, nucleolii nu mai sunt vizibili. Cromatina este dispusa in
gramezi dense si neregulate, colorate violet negricios. Citoplasma
este intens bazofila ("cea mai albastra celula"), datorita
bogatiei de poliribozomi si proteine. Reprezinta in medie
16% (10-25%) din elementele seriei rosii si 2-4% din totalul
eritroblastilor.
Eritroblastul policromatofil este de talie mai mica (10-12 m),
frecventa este de aproximativ 50% din elementele seriei rosii.
Nucleul este rotund, condensat, raportul nucleu/citoplasma este egal sau
chiar in favoarea citoplasmei. Cromatina nucleara este mai
condensata, aspect de "spita de roata", dar nu atat de
evidenta ca la plasmocit. Citoplasma este mai bazofila la periferie,
apoi spre periferie o tenta mai portocalie, ca expresie a sintezei Hb.
Reprezinta 50% din elementele seriei rosii si 10-20% din totalul
eritroblastilor.
Eritroblastul oxifil (ortocromatic) este cel mai mic precursor al seriei
rosii (8-10 m), frecventa pe frotiu fiind in
medie de 32% din seria rosie. Nucleul se reduce treptat, devine excentric,
picnotic, intens colorat (tahicromatic). Citoplasma este abundenta,
oxifila (portocalie) datorita sintezei Hb si scaderii
ribonucleoproteinelor. Maturarea sa consta in eliminarea nucleului
(expulzia sa), ce se realizeaza la nivelul MO cu ajutorul macrofagelor.
Reprezinta 10-20% din totalul eritroblastilor.
Reticulocitul este de talie mai mare decat
eritrocitul (9-10 m). Durata de viata este de
72 ore, din care 48 ore le petrece in MO si 24 ore in sangele periferic.
Reticulocitul prezinta o retea filamentoasa alcatuita
din ARNm si ribozomi ce le permite o activitate metabolica
intensa fata de eritrocit (continua sinteza Hb pana la
saturarea eritrocitelor; confera celulei o activitate respiratorie-ciclul
Krebs; permite miscari de diapedeza, cu ajutorul carora pot
trece din spatiul medular in sange). Trecerea reticulocitelor din MO in
sangele periferic se face prin traversarea peretilor sinusurilor medulare,
proces denumit diapedeza.
Reteaua caracteristica
reticulocitului se evidentiaza prin coloratii supravitale
(albastru crezil briliant, rosu neutru, albastru de metilen).
Odata cu maturarea, aceasta
retea dispare si reticulocitul devine hematie.
Valoarea normala a
reticulocitelor este de 0,5-1,5, maxim 2% (la 100 eritrocite) sau
5-150/00 (la 1000 eritrocite); in valoare absoluta
20000-80000/mm3. La nou-nascut in primele zile valorile
ajung la 600/00 (reticulocitoza
fiziologica a nou-nascutului), apoi dupa prima
saptamana de viata valorile scad rapid, ajungand in
luna a-3-a la 50/00.
Cresterea numarului de
reticulocite poate avea o valoare prognostica buna (in tratamentul
anemiilor aregenerative) sau o valoare diagnostica (in tratamentul
anemiilor hemolitice). Scaderea numarului de reticulocite apare in aplazia
medulara de orice cauza, in blocajul de maturare a
eritroblastilor si in transformarea sau invazia maligna a
maduvei (cancere, hemopatii).
In anemiile prin carenta de
factori necesari eritropoiezei (Fe, vitamina B12, acid folic,
proteine), administrarea factorului carential determina
cresterea numarului de reticulocite in sangele periferic "criza reticulocitara", ceea
ce indica ca maduva este reactiva, cu potential bun
hematoformator (cauza este extramedulara) si tratamentul anemic este
eficient.
In anemiile din aplazia
medulara, tratamentul cu factori necesari eritropoiezei determina
cresteri nesemnificative ale reticulocitelor (maduva este
hipo/areactiva).
REGLAREA ERITROPOIEZEI
Reglarea eritropoiezei se
realizeaza prin mecanism de tip feed-back pozitiv asupra celulelor
nediferentiate, semnalele declansatoare fiind generate in
tesuturi, ca urmare a variatiilor aportului de oxigen.
Reglarea eritropoiezei si adaptarea ei permanenta in functie de
necesitatile de oxigenare ale tesuturilor se face prin mecanisme
complexe umorale si nervoase.
Reglarea umorala Factorul
esential care determina cresterea sau scaderea ratei
productiei medulare de eritrocite este concentratia oxigenului in
sangele arterial. Scaderea oxigenului duce la hipoxie tisulara,
intalnita la altitudine, in starile de alcaloza, in
insuficienta cardiaca, hemoragii repetate, intoxicatii cu oxid
de carbon, distructii medulare, etc.
Reglarea umorala a eritropoiezei
se realizeaza prin:
eritropoietina (vezi Reglarea
hematopoiezei")
FSC-GEMM (IL-3)
Alte interleukine: IL-4, IL-9
in mai mica masura prin alti
hormoni:
insulina,
androgenii, tiroidienii, adrenalina stimuleaza eritropoieza;
cortizolul,
estrogenii o inhiba
Reglarea nervoasa a eritropoiezei se realizeaza
direct (reflex) prin intermediul unor formatiuni vegetative sau indirect
(neuro-umoral) prin modificarea secretiei de eritropoietina.
Mecanismul reflex este declansat
de stimularea chemoreceptorilor sinocarotidieni, ca urmare a scaderii
presiunii partiale a oxigenului din sangele arterial. Impulsurile aferente
sunt conduse prin fibrele aferente ale nervilor Hering (ramuri ale nervilor
glosofaringieni) la centrii nervosi din substanta reticulata
bulbara si hipotalamusul posterior, de unde pornesc impulsuri efectoare
conduse prin fibre care trec prin maduva in nervii splahnici si
apoi ajung la maduva hematogena. Activarea simpatoadrenergica
are ca efect cresterea numarului de eritrocite si reticulocite,
datorita mobilizarii sangelui din organele de depozit si
stimularea citodiabazei
In productia si
functionalitatea normala a eritrocitelor un rol important revine
si vitaminelor si oligoelementelor, care intervin ca factori de crestere
sau de maturare:
vitamina B12 (cobalamina) si
acidul folic = factori de maturare. Deficitul induce anemia megaloblastica
Biermer (anemia hipercroma) caracterizata prin: numar
scazut de eritrocite, Hb scazuta, Ht scazut, VEM crescut,
HEM scazut, CHEM normal.
Fierul = factor de crestere,
necesar pentru sinteza hemului din hemoglobina. Deficitul de Fe
determina anemia microcitara, hipocroma, feripriva,
caracterizata prin: numar scazut de eritrocite, Hb
scazuta, Ht scazut, VEM scazut, HEM scazuta, CHEM
scazuta, sideremia (Fe circulant sau concentratia Fe seric)
scazuta.
Alti factori: vitamina B6,
vitamina C; cuprul, cobaltul, zincul, proteinele.
Eritrocitul. (hematia)
este elementul final al seriei rosii. Este celula purtatoare de
pigment respirator - hemoglobina. Specializarea sa in transportul de gaze,
avand astfel o functie vitala pentru organism, inseamna si
renuntarea la o structura si proprietati normale.
Isi pierde nucleul, organitele celulare si o mare parte din
echipamentul enzimatic, ceea ce duce la reducerea duratei de viata la
aproximativ 120 de zile, deoarece enzimele degradate si componentele
structurale uzate nu mai pot fi inlocuite prin sinteza.
Eritrocitul parcurge
cca. 300 Km, pe acest drum fiind supus agresiunilor mecanice in capilare - mai
ales in cele splenice - pe care le poate traversa doar cu mari modificari
de forma si volum, dar si la agresiuni biochimice, atat din
exteriorul celulei, cat si din partea continutului - oxigenul
transportat, care actioneaza ca un puternic agent oxidant.
La om si la
mamifere, eritrocitele (E) sunt celule incomplete, anucleate, incapabile
sa mai sintetizeze proteine si din acest motiv nu-si pot
pastra structura mult timp. La reptile, pasari si
batracieni E sunt ovalare, cu nucleu.
Denumirea
eritrocitului vine de la "eritos" (rosu) si de la "citos"
(celula), denumire ce a fost contestata datorita absentei
nucleului. Metodele moderne de histochimie, citochimie si microscopie
electronica impreuna cu argumente de filo si ontogenie au
consacrat recunoasterea E ca o celula de sine statatoare,
dar cu o activitate metabolica mai restransa si durata de
viata limitata.
Rolul principal
este de a transporta O2 de la plamani la tesuturi si
a unei parti din CO2 de la tesuturi la plamani.
In plus, prin sistemele tampon pe care le contine, E participa la
mentinerea echilibrului acido - bazic.
Constante eritrocitare directe
Forma: -
disc biconcav, cu margini rotunjite, cu periferia mai colorata si
centrul mai palid (figura 4.3);
- pe sectiune apare ca un piscot sau
ca o haltera. Aceasta forma este o adaptare la functia de
transport a O2 - asigura
eritrocitului un maxim de suprafata de schimb pentru volumul dat, cu
o cincime mai mare decat forma sferica la acelasi volum (plansa
1).
Culoarea: -
pe frotiul MGG este roz, mai intensa la periferie, mai palida in
centru unde sunt mai subtiri.
Diametrul: -
7,2 μ, cu limite intre 6,68 - 7,72 μ
Variatii:
- la nastere exista macrocitoza fiziologica: 8 - 9 μ
si curba Price - Jones este deviata la dreapta;
- diametrul scade apoi pana la 4-5 luni,
ajungand intre 5-6 μ. Scaderea diametrului este insotita
si de scaderea numarului de E (este inlocuita Hb F cu Hb
A). Spre sfarsitul primei decade de viata ajunge la valori
normale.
Densitatea: 1,090 - 1,093. Cum densitatea
sangelui este de 1,050 - 1,070, in vitro E
se depun la fundul eprubetei - sedimenteaza.
pH:
Cantitatea de Hb: 14 - 16g /dl. Scaderea sub 11g% poarta numele de anemie.
Numarul de E: in medie este de 4,5 - 5 milioane / mm3.
Variatii
fiziologice ale numarului:
▪ legate de sex: la barbati
numarul este mai mare numai dupa pubertate datorita hormonilor
androgeni, care stimuleaza eritropoieza: 4,9 ± 0,7 milioane/mm3,
iar la femei este 4,3 ± 0,6 milioane /mm3 (estrogenii deprima
eritropoieza);
▪ varsta: nou-nascutul are un numar mai mare (datorita
transfuziei cu sange matern prin cordonul ombilical). Dupa 6 - 12
saptamani numarul scade catre valoarea de anemie
datorita inlocuirii Hb F cu Hb A. Dupa 6 luni numarul incepe
sa creasca ajungand la valoarea de la adult la aprox. 12 ani. La
batrani scade (chiar la valori de anemie) prin inlocuirea maduvei
rosii hematogene cu maduva cenusie, fara
potential hematogen;
▪ altitudine: prin scaderea pO2
se instaleaza hipoxia
tisulara, care induce poliglobulie
fiziologica de altitudine. Adaptarea rapida la altitudine
(ascensiuni montane intr-un timp scurt) se realizeaza in primul rand prin
acumularea intraeritrocitara de 2,3 - DPG, urmata de cresterea
eliberarii O2 la tesuturi si apoi prin contractia splinei,
care elimina in circulatie hematiile depozitate. Cresterea numarului de hematii modifica
proprietatile reologice ale sangelui - creste vascozitatea
si implicit rezistenta vasculara periferica pe care cordul
o va invinge prin cresterea TA; de aceea expunerea la altitudine (peste
900m) este contraindicata persoanelor hipertensive, cu afectiuni
cardiace, sau CPC (cord pulmonar cronic), la care poate aparea un edem
pulmonar acut. La expunerea de durata la altitudine (hipobarism prelungit)
adaptarea se face prin secretie crescuta de eritropoietina (EPO)
a carei sinteza la nivel renal este stimulata de hipoxie. EPO
actioneaza la nivelul maduvei osoase hematogene (MOH) stimuland
producerea de E, care vor aparea in sangele periferic dupa 7-10 zile,
timpul necesar diferentierii si maturarii precursorilor.
Persoanele care locuiesc permanent la inaltimi mari, de exemplu
locuitorii din Tibet sau muntii Anzi (peste 5000m) au un numar de E
permanent crescut, care ajunge la 7 -
7,5 milioane E / mm3.
Cresterea
presiunii atmosferice, situatie mai rar intalnita - munci la adancimi
mari (scafandri, mineri ) determina scaderea numarului de
eritrocite..
▪ efortul fizic:
◦
moderat: creste numarul eritrocitelor prin pierderea de apa in
interstitiu si hemoconcentratie, prin splenocontractie (Ht
splenic este de 70 - 80%) si prin stimularea citodiabazei.
◦
intens: scade numarul de E, deoarece apare fenomenul de eritroliza,
in principal prin cresterea vitezei de circulatie a sangelui si
traumatizarea E in timpul traversarii vaselor. Poate aparea astfel hemoglobinuria paroxistica de efort
(cunoscuta si ca hemoglobinuria de mars).
▪ activitatea intelectuala, stresul,
emotiile: creste numarul de E, ca urmare a secretiei
crescute de adrenalina si cortisol.
▪ cresterea temperaturii:
creste nr. E prin splenocontractie
▪ somnul: scade numarul
▪ digestia,
starea de repaus sau de activitate, modificarile de pozitie induc
variatii mici (intre 5 - 10%) ale numarului de E ceea ce nu
justifica postul alimentar si conditiile riguroase de recoltare
a probelor de sange (doar in dozarea
glicemiei sunt recomandate restrictiile).
Variatii
patologice
- cresterea
numarului peste 6 - 6,5milioane./mm3 poarta numele de poliglobulie
sau policitemie. Aceasta poate fi secundara altor boli care determina
hipoxie (CPC, patologie cardiaca, etc.) sau primara - policitemia
Vera > 12 milioane E / mm3 (proliferarea maligna a seriei rosii).
- scaderea sub 3,5
- 4 milioane / mm3 poarta numele de anemie.
Volumul elementelor figurate exprimat procentual din volumul total de sange = hematocrit (Ht) = volum globular.
Valoarea normala: 40 - 45% ± 5%. Germanii exprima si in
l de E/l sange: la barbati 0,40 - 0,48 l / l, iar la femei 0,36 - 0,42 l / l.
Determinarea Ht si a Hb sunt cele mai
fidele si cele mai utilizate investigatii de laborator pentru
diagnosticul anemiilor si al poliglobuliilor.
Variatii
fiziologice ale Ht
- functie de sex: Barbati: 46 ± 5%; Femei:
40 ± 5%;
- varsta: n.n.- 56%; 6 luni - 36%; 1 an - 32%; 4 ani - 37%; 12
ani 40%;
- efort fizic: creste prin splenocontractie si prin
pierderea de apa, ca urmare a cresterii permeabilitatii
capilare - hemoconcentatie;
- altitudine: Ht crescut prin poliglobulie determinata de
hipoxia tisulara - scade pO2 in
aerul inspirat;
- sarcina: Ht scazut prin hemodilutie - creste
volumul plasmatic ca urmare a retentiei de Na+ si
apa, datorate excesului de estrogeni si progesteron;
- teritoriul vascular: sange arterial: 45%; sange venos 47%; sange
splenic: 70 - 80%
Constantele
eritrocitare enumerate sunt constante directe (masurabile direct). Pe baza
lor se calculeaza constantele indirecte sau derivate. Acest calcul se face
cu ajutorul unor formule matematice, valorile depinzand de exactitatea
determinarii constantelor directe.
Constane eritrocitare indirecte (derivate)
VEM = volumul eritrocitar
mediu = (Ht  10) / nr. E = 80 -
94 μ3 sau fentolitri (≈ 87 μ3 ).
In engleza: MCV = mean volume of one RBC (Red Blood Cell). Pentru
numarul de E se iau primele 2 cifre cu virgula
10) / nr. E = 80 -
94 μ3 sau fentolitri (≈ 87 μ3 ).
In engleza: MCV = mean volume of one RBC (Red Blood Cell). Pentru
numarul de E se iau primele 2 cifre cu virgula
Cresterea peste 94 μ3 - anemii macrocitare; peste 120 μ3
- anemii megalocitare;
Scaderea sub 80 μ3 - anemii microcitare
HEM = cantitatea de Hb
eritrocitara medie
= Hb (g%) 10 / nr. E = 25 - 32 μμg (pg) ≈ 29
μμg (pg). In engleza: MCH = mean HB mass/RBC
Cresterea peste 32 μμg (pg) - anemie Biermer, unde cantitatea
absoluta de Hb in E este mai mare, datorita VEM crescut.
Scaderea sub 25 μμg (pg) - in anemii hipocrome, microcitare, unde
cantitatea absoluta de Hb este scazuta datorita volumului scazut si
slabei incarcari cu Hb a microcitelor
CHEM = concentratia de Hb eritrocitara medie din 100 ml de
masa eritrocitara =
Hb 100 / Ht = 32 - 36 g Hb / 100ml masa
eritrocitara. In engleza: MCHC = mean HB conc. Ib RBCs.
Este cea mai
precisa constanta, deoarece metodele de determinare a Hb si Ht
au erori minime.
Scaderea sub 32% presupune o incarcare redusa cu Hb - anemii
hipocrome, microcitare
Cresterea peste 36% nu este posibila (cresterea volumului E
influenteaza si cresterea Ht, valoarea raportului ramanand
constanta). In anemiile macrocitare si megaloblastice CHEM este
normala, iar hipercromia este falsa.
STRUCTURA ERITROCITULUI
Structura eritrocitului
este adaptata functiei de transportor de gaze si
conditiilor particulare in care circula prin diversele teritorii
vasculare. Pentru a se putea deplasa, adaptarea esentiala consta
in elasticitatea si deformabilitatea mare pe care o are aceasta
celula. Adaptandu-se insa la mediul de circulatie, eritrocitul
sufera modificari importante fata de alte celule, atat in
structura membranei cat si a citoplasmei.
Membrana. Contine
proteine, lipide si hidrati de C, care sunt de obicei asociati
cu proteine.
Prin bariera pe care o
realizeaza intre citosol si mediul plasmatic, membrana
protejeaza Hb eritrocitara. Daca s-ar afla libera in
plasma, Hb ar crea o presiune coloid-osmotica suplimentara
presiunilor deja existente, ducand la o crestere a presiunii arteriale la
valori incompatibile cu viata.
Studiul membranei
eritrocitare s-a realizat prin obtinerea stromelor sau "fantomelor"
eritrocitare rezultate in urma golirii eritrocitelor de continut,
dupa care componentele membranare au fost analizate separat. Acest model a
servit pentru intelegerea structurii si a altor membrane decat cea
eritrocitara.
Membrana eritrocitului
este formata din 3 straturi: superficial, mijlociu si intern (figura
4.1).
Stratul superficial. Are grosimea de 20Ǻ si este format din lanturi de
hidrati de C, singuri sau in asociere cu proteinele, formand glicoproteine
(GP). Clasic se numeste glicolema. O parte dintre oligozaharide se
termina cu acid sialic, care imprima incarcatura
electrica negativa a exteriorului celulei.
GP de suprafata
au rol de antigene specifice eritrocitare: antigenele grupelor si
subgrupelor sanguine, de receptori pentru virusuri, paraziti, lecitine
vegetale si rol in recunoasterea celulara si adezivitate.
Stratul mijlociu este membrana propriu-zisa, cu o structura lipidica bistratificata.
Straturile lipidice ancoreaza la suprafata polizaharide si
sunt strabatute de proteine tubulare numite proteine integrate, unele
dintre acestea - glicoforinele A, B si C, proteina benzii 3 - fiind
adevarate canale de transport. Lipidele membranare provin din plasma.
Stratul lipidic este
foarte mobil, fluid si se onduleaza ca un strat de petrol la
suprafata valurilor, conferind eritrocitului suplete si o mare
capacitate de deformare Rolul esential in fluiditatea membranei il are
colesterolul, care alaturi de spectrina, proteina specifica
eritrocitului, sustine capacitatea de a suferi deformari reversibile
fara variatii ale volumului sau suprafetei, avand rol
important si in determinarea dimensiunilor celulei. Scaderea
cantitatii de colesterol reduce suprafata eritrocitului, care
devine sferocit, iar excesul mareste suprafata, aparand
hematii in tinta.
Stratul intern
alcatuieste scheletul membranei, structura de suport a membranei.
Este format dintr-un numar mare de proteine fibrilare (separate prin
electroforeza), care formeaza o retea de suport. O parte a
acestor proteine au primit ca denumire un numar, corespunzator
pozitiei pe care au ocupat-o in migrarea electroforetica. Ele sunt
ancorate de bistratul lipidic prin intermediul unor proteine integrate.
Exista 2 tipuri de proteine:
- proteine integrate: proteina benzii 3
(schimbator anionic), aquaforinele (canale pentru apa),
glicoforinele. Aceste proteine se gasesc in citoschelet si
strabat stratul mijlociu fosfolipidic. La suprafata celulelor ele
contribuie la formarea antigenelor si receptorilor membranari. In
celula se leaga de spectrina, fie direct fie prin intermediul
altor proteine. (de exemplu ankirina leaga proteina benzii 3 cu
reteaua de fibrina).
- proteine periferice, fibrilare; se
gasesc pe suprafata citoplasmatica a bistratului lipidic: spectrina, actina, ankirina, proteina
benzii 4.1, tropomiozina, tropomodulina etc..
Spectrina este o proteina care se
gaseste numai in eritrocit. Are aspect fibrilar, formand retele
tridimensionale (plasa) care sprijina stratul lipidic.
Legaturile dintre fibrele de spectrina (nodurile) se fac prin
intermediul unor proteine globulare - actina, proteina benzii 5, a benzii 4.1,
a benzii 2, etc.. Interactiunile verticale se realizeaza cu stratul
lipidic, prin intermediul proteinelor integrate - proteina benzii 3
si glicoforina A.
Structura
defectuoasa sau absenta acestor proteine duce la alterari ale
membranei, consecinta fiind incapacitatea de a-si pastra forma
(devin sferocite sau ovalocite) si proprietatile, mai ales
deformabilitatea. Astfel de defecte membranare determina liza, moartea
precoce a eritrocitelor, ceea ce va duce la instalarea unei anemii hemolitice.
Glicoforinele
A, B,C ,D
prezinta ramificatii terminate cu oligozaharide. O parte dintre
oligozaharide se termina cu acid sialic, care imprima
incarcatura electrica negativa a exteriorului
celulei. Datorita acestei incarcaturi electrice intre eritrocite
se manifesta forte de respingere, care intarzie aglutinarea si
sedimentarea, permitand mentinerea in suspensie a eritrocitelor.
Scaderea cantitatii de acid sialic la eritrocitele senescente
este unul dintre factorii care favorizeaza captarea splenica.
Citosolul - mediul intern al eritrocitului sau stroma. Contine 60% apa, 33-35%
Hb si 5-7% alte substante. Este o suspensie apoasa in care se
gasesc : macromolecule (Hb si enzime); substante organice cu
molecula mica (glucoza, glutation, ATP); compusi metalici
si metaloizi (P, S, Cu, Zn etc.)
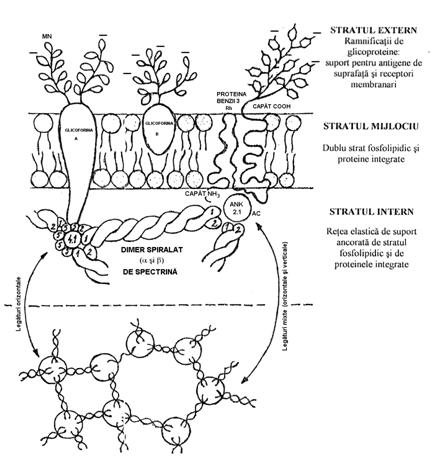
Figura nr. 4.1. Structura schematizata a membranei eritrocitare -
sectiune frontala (dupa A. Valcu, 1997).
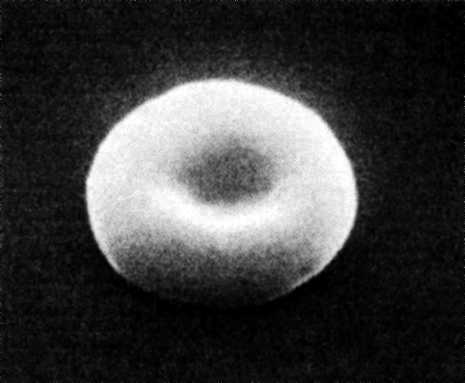
Figura nr. 4.3 Eritocitul normal. Microfotografie prin scanare cu
electroni. Se observa forma de disc biconcav, si grosimea mai
mica in centru, ceea ce face sa para mai palide in mijloc
si mai intense la periferie (dupa Stanley L Schrier - 2000).
METABOLISMUL ERITROCITULUI
Pentru a-si
indeplini functiile si pentru a-si pastra integritatea, E
consuma energie furnizata de ATP. Acesta rezulta din
metabolizarea aproape in exclusivitate a glucozei. S-a constatat insa
ca eritrocitul mai poate metaboliza si galactoza, fructoza
si manoza.
Deoarece nu are depozite de glicogen (decat urme), E este total
dependent de glucoza din sange Intrarea glucozei in E se face prin difuziune
facilitata, neavand nevoie de insulina (adaptare pentru a proteja E,
a carui functie este vitala pentru organism).
In orice metabolism
celular, sistemele enzimatice sunt esentiale. Din pacate, E a pierdut
in procesul de maturare organitele (nucleul, mitocondriile, ribozomii, aparatul
Golgi, lizozomii) si enzimele implicate in sintezele proteice. Enzimele
"mostenite", degradate pe parcursul vietii eritrocitare, nu pot fi
inlocuite si celula va muri, fiind lipsita de energie si
potential reducator.
Indeplinirea
functiilor se poate realiza, deoarece E adult paraseste MOH
inzestrat cu un echipament enzimatic complet, necesar degradarii glucozei
si deci obtinerii de energie, precum si cu enzime
reducatoare necesare protejarii Hb.
Spre deosebire de alte
celule, E nu contine enzimele ciclului Krebs si este singura
celula care functioneaza fara sistemul citocromic.
Mai
pastreaza: enzimele ciclului Embden - Mayerhof: hexokinaza,
triozo-fosfat-izomeraza, fosfoglicerokinaza, piruvat-kinaza; enzimele ciclului
pentozo-fosfatilor: glucozo-6-fosfat-dehidrogenaza; enzime
reducatoare: methemoglobin-reductaza care asigura transformarea
majoritatii metHb (forma oxidata, inactiva a Hb) in Hb
activa, capabila sa lege O2.
Degradarea glucozei si furnizarea energiei. Glucoza este metabolizata prin:
1). Calea Embden - Mayerhof, care are o pondere de 90 - 95%
in degradare. Un mol de glucoza furnizeaza 2 moli ATP folositi
pentru energie in glicoliza si 2 moli ATP furnizati sistemelor
celulare (ciclul Krebs furnizeaza 38 de moli de ATP), deci randamentul
acestei cai este mic. Este suficient insa, deoarece E are un consum
energetic redus. Desi cantaresc 5% din greutatea organismului,
eritrocitele consuma doar 1% din metabolismul bazal (figura 4.4)
In urma reactiilor
acestei cai, H+ rezultati sunt acceptati de NAD+
care se transforma in NADH, acesta reprezentand furnizorul de H+
utilizati la reducerea metHb. Produsii finali ai acestei cai,
piruvatul (care in alte celule intra in ciclul Krebs) si acidul
lactic, difuzeaza din E si sunt metabolizate in alte tesuturi.
Calea Embden-Mayerhof este importanta pentru:
- sinteza de
ATP. Acesta este cel mai important intermediar energetic in E. Este utilizat ca
acumulator de energie necesara mentinerii formei si
flexibilitatii membranei, functionarii normale a pompelor
ionice si substituie, la nevoie, 2,3-DPG in procesul de disociere a oxiHb;
- generarea
de NAD+ (nicotinamid - adenindinucleotid) si forma sa
redusa NADH - important sistem redox al celulei. NADH este esential
pentru mentinerea Fe2+ din hem in stare redusa, proces
catalizat de NADH-methemoglobinreductaza.
2). Suntul
Rappaport - Luebering este o cale derivata, secundara a caii Embden-Mayerhof,
specifica E prin care se sintetizeaza 2,3 difosfogliceratul
(2,3-DPG). Roluri:
-
conserva excesul de energie rezultat din calea Embden-Mayerhof
- intervine
in transportul transmembranar al K+
- in
conversia metHb in oxiHb (acumularea sa genereaza NADH care activeaza
enzima NADH- methemoglobinreductaza)
-
modifica curba de disociere a oxiHb: prin legarea sa de Hb favorizeaza
cedarea O2 catre tesuturi, iar prin desprinderea sa de pe
Hb favorizeaza legarea labila a O2 la Hb.
Primul si cel mai
rapid mecanism de adaptare la altitudine este acumularea intraeritrocitara
a 2,3-DFG, urmata de cresterea eliberarii O2 la
tesuturi (acest mecanism este prezent si in prima etapa a
anemiilor). Urmeaza apoi contractia splinei si stimularea prin
EPO.
3).Ciclul pentozelor /
suntul pentozo-monofosfatilor este o cale aeroba de degradare a
glucozei. Are o pondere de 5-10%, putand ajunge pana la 20%, in
functie de necesitati. Are aport energetic scazut, dar
reprezinta singura cale de sinteza intraeritrocitara a NADPH+
- sistem reducator de rezerva, care protejeaza Hb de
agentii oxidanti. Pentru fiecare mol de glucoza se produc 2 moli
de NADPH.
NADPH este necesar
pentru mentinerea in echilibru a sistemului oxidoreducator glutation
redus/glutation oxidat care protejeaza Hb de agentii oxidanti
(partea proteica a Hb - globina).
Glutationul este
sintetizat intraeritrocitar. Este un tripeptid (acid glutamic-glicocol-cisteina).
El protejeaza gruparile sulfhidrice (-SH) ale componentei proteice a
Hb - globina de oxidare, impiedicand formarea metHb si a corpilor Heinz.
In procesul de reducere a gruparilor SH oxidate, glutationul se
oxideaza, transformandu-se in glutation oxidat. Acum intervine enzima
glutation reductaza care reduce glutationul oxidat, refacand
concentratia de glutation.
Oxidarea Hb este o reactie spontana, normala, care are
loc permanent in hematie, tocmai datorita O2 transportat, sau sub actiunea
peroxidului de H (H2O2) si a altor radicali oxizi
liberi (O2- , OH) care iau nastere spontan sau
dupa administrarea unor medicamente cu efecte oxidante. H2O2
este foarte nociv pentru orice celula si poate oxida toate
componentele proteice - Hb, enzimele membranare sau citosolice).
In concluzie: energia obtinuta prin cele
3 cai si compusii biochimici rezultati sunt folositi
astfel:
- pentru mentinerea
integritatii membranei si constantei mediului intern al
celulei, adica pentru functionarea in principal a pompelor de
Na+-K+, Mg+, Ca2+, etc.;
- pentru mentinerea
Fe feros din Hb (Fe2+), O2 legandu-se reversibil
numai de acesta. Formarea Fe3+ (fier feric) produce metHb care este
un produs inactiv, incapabil sa mai cedeze O2 . Acest proces se
produce permanent in E, iar reducerea Fe3+ la Fe2+ este
un proces foarte important pentru E, care este asigurat de sistemul
reducator principal: NADH-Hbreductaza si de sistemul reducator
accesor (cu pondere mai mica) - NADPH-hemoglobinreductaza;
pentru protejarea globinei si a membranei impotriva oxidarii,
care este asigurata de sistemul glutation redus / glutation oxidat;
- pentru formarea
2,3-DFG asigurata de suntul Rappaport - Lüebering.
Proprietatile
fizico-chimice ale eritrocitului
1.
Elasticitatea sau deformabilitatea
Este capacitatea eritrocitelor integre morfologic si
functional de a-si adapta forma la fortele permanente de
forfecare care se exercita atat din interiorul cat si din exteriorul
lor. Abilitatea de a-si modifica forma permite eritrocitelor sa strabata
sectorul microcirculatiei care cuprinde capilare cu f<3m, asigurand transportul gazelor respiratorii catre si de la
celule, facilitand in acelasi timp si transportul apei din
spatiile interstitiale in sange (figura 4.5.).
Factorii care asigura deformabilitatea
▪ Forma de disc biconcav
care asigura eritrocitelor un maximum de suprafata de schimb
pentru volumul dat. Mentinerea acestei forme se face cu o mare
cheltuiala energetica, energia furnizata de desfacerea
legaturilor fosfat macroergice ale APT-ului fiind folosita pentru
mentinerea in activitate a pompelor de ioni, care trebuie sa conserve
un gradient de concentratie optima pentru principalii ioni
implicati in schimburile transmembranare: Na+ K+, Ca2*,
Cl_, HCO3-.
A
B
Figura
nr. 4.5. Circulatia eritrocitelor in capilarele cu diametrul mai mare
de 5 mm (A) si in
capilarele cu diametrul intre 3-5 mm
(B) (dupa Valcu, A., 1996)
▪ Fluiditatea
continutului eritrocitar sau vascozitate interna
infuentata in special de continutul in Hb. Existenta unor
Hb anormale , mai ales a Hb S, care cristalizeaza in stare redusa sau
deshidratarea severa a eritrocitului au drept consecinta
scaderea fluiditatii si implicit a
deformabilitatii.
▪ Compozitiei membranei bogata in fosfolipide (lecitina, lizolecitina) si colesterol, cu dispozitie
asimetrica.
▪ Arhitectura
citoscheletului eritrocitar conferita de proteine ca spectrina,
actina, proteina benzii 4.1, ankirina, proteina benzii 3, glicoforine.
Alterarea mitogena sau biochimica a proteinelor scheletale duce la
instabilitatea interactiunlor specifice, urmata de pierderea unor
fragmente membranare, de scaderea suprafetei si a volumului, de
micsorarea deformabilitatii, de augmentarea captarii
splenice si a riscului de eritroliza precoce (Mohandas, N., 2001).
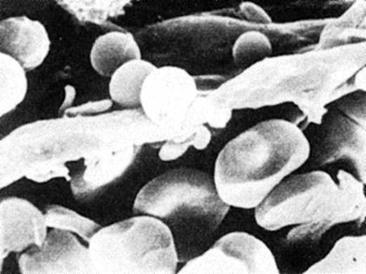
Figura nr. 4.6. Eritocite in sinusoidele
splenice. Microfotografie prin scanare cu electroni. Pentru a trece bariera
sinusoidala eritrocitele se deformeaza; sferocitele sunt incapabile sa traverseze
aceasta bariera (dupa
Mohandas Narla - 2001).
|
|
2. Dispunerea in
fisicuri
este proprietatea hematiilor de a adera intre ele cu formarea de fisicuri,
sub aceasta forma circuland in capilarele cu F>10m (fig. 4.5. A). In
aceste vase, sangele curge intr-un mod particular, grupurile de
fisicuri-trenuri fiind separate de boluri de plasma (curgere
start-stop). Un tren de eritrocite il izbeste pe cel din fata sa
proiectandu-1 inainte, in timp ce el este impins de trenul din spatele
sau. Dispunerea in fisicuri a hematiilor este reversibila
si depinde atat de calitatile lor, cat mai ales de
existenta in plasma a unor proteine asimetrice care pot augmenta sau
deprima aceasta capacitate (albumine, globuline, fibrinogen). Marimea
fisicurilor influenteaza viteza de sedimentare a hematiilor in vitro.
3. Stabilitatea in
suspensie a eritrocitelor Diferenta de densitate dintre
plasma -1027 si eritrocite -1093, ar trebuii sa duca,
conform legii gravitatiei la sedimentarea rapida a hematiilor.
Totusi, 'in vivo'
hematiile sunt mentinute in suspensie, iar in vitro, sedimentarea se produce lent.
Dispersia uniforma a eritrocitelor in plasma este
mentinuta de mai multi factori:
▪ Fluxul sanguin este unul dintre factorii cei mai importanti,
evidentiat prin sedimentarea rapida a hematiilor la oprirea
circulatiei printr-un sector vascular.
▪ Factori plasmatici:
intre eritrocite, care au o incarcatura predominant
negativa conferita de acidul sialic (dispus pe suprafata
celulei), se manifesta forte de respingere electrostatica, care
fac posibila mentinerea stabilitatii in suspensie.
Prezenta in plasma a unor proteine cum sunt fibrinogenul, Ig A, G, M,
a1-glicoproteinele, a2 macroglobulinele,
purtatoare ale unor incarcaturi electrice predominant pozitive,
neutralizeaza sarcinile negative, initiale ale eritrocitelor si
favorizeaza formarea agregatelor eritrocitare. Asa se explica
cresterea VSH-ului in starile in care creste concentratia
globulinelor - infectii acute si cronice, boli hepatice, nefroze,
etc. sau a fibrinogenului - sarcina, menstruatie, batrani, etc..
Alte proteine plasmatice, albuminele, cu o incarcatura
predominant negativa (data de anionii pe care-i fixeaza) cresc
fortele de respingere manifestate intre eritrocite si sedimentarea se
produce mai greu (VSH scazuta).
▪ Factori eritrocitari - marimea si densitatea agregatelor
eritrocitare este influentata de numarul, morfologia si
continutul in Hb al hematiilor.
VSH este invers proportionala
cu hematocritul. Astfel:
- cresterea numarului de
hematii - poliglobulie se insoteste de scaderea VSH (surplus de
sarcini electrice negative, forte de respingere mai mari, stabilitate a
sistemului).
- scaderea numarului de hematii,
asociata sau nu cu scaderea concentratiei proteinelor plasmatice
creste VSH (in aceasta situatie, la deficitul de sarcini
negative se adauga si scaderea vascozitatii sangelui).
- anizocitoza (variatiile de marime)
favorizeaza formarea de rulouri neuniforme si o sedimentare in
straturi.
- modificarile de forma - platicitele,
sferocitoza, acantocitoza si deficitul de Hb impiedica formarea de
agregate, incetinind viteza de sedimentare.
▪ Factori fizico-chimici:
- temperatura crescuta creste VSH;
- inclinarea tuburilor la 45° (metoda
Fuente-Hita) creste valoarea de 4 ori;
- inaltimea coloanei- tuburile
trebuie sa aiba aceeasi lungime: 20 cm, cu cat creste
inaltimea coloanei, creste acceleratia, creste viteza;
- natura si concentratia
anticoagulantului. O cantitate mai mare de anticoagulant produce dilutie
si scaderea VSH.
Metoda de determinare a VSH,
cea mai des folosita in clinica este metoda Westergreen. Valori normale:
B: 7-9mm/h; 10-12mm/2h; F:
8-10mm/h; 12-16mm/2h; nou - nascuti : 1 - 2 mm/h (numar mare de
eritrocite)
Importanta
determinarii:
VSH este un indicator clinic nespecific al starii de boala, indicand
un raspuns imunologic in desfasurare, fara a preciza
natura acestuia, localizarea sau gravitatea bolii care l-a indus.
Ca element de urmarire in dinamica a unei patologii,
cresterea VSH semnifica acutizarea bolii, iar scaderea -
regresiunea bolii.
In bolile acute, VSH crescuta semnifica o evolutie
favorabila a raspunsului imunologic, iar absenta cresterii,
intr-un context clinic evident de patologie infectioasa, se
coreleaza cu un sistem imunitar areactiv. In bolile cronice,
cresterea VSH sugereaza activarea procesului inflamator.
4. Scintilatia. Este
proprietatea eritrocitelor normale de a scanteia cand sunt examinate la
microscop; se datoreaza miscarilor vibratorii care
influenteaza indicii de refractie. Este absenta daca
eritrocitul este rigid, precum si in cazul eritrocitelor fixate pe
lama.
5. Rezistenta
globulara
In conditii normale, hematiile integre morfologic si
functional pot rezista la agresiunea unor factori fizici, chimici si
biologici fara a liza.
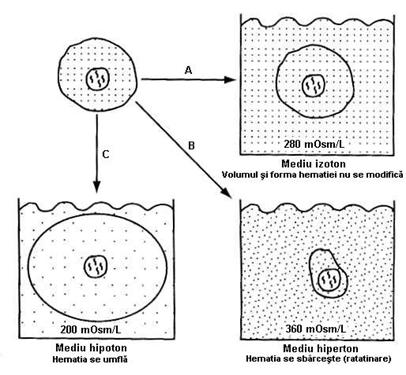
Figura 4.7 - Efectele unei solutii izotonice (A), hipertonice (B)
si hipotonice (C) asupra volumului si formei celulelor (hematia)
In vitro, rezistenta globulara se
apreciaza prin expunerea hematiilor la agenti fizici - ultrasunete,
incalzire; chimici - acizi, baze, cloroform, alcool; biologici - anticorpi
antieritrocitari sau fizico-chimici - scaderea presiunii osmotice a
mediului de suspensie. In cazul determinarii rezistentei osmotice se
realizeaza un mediu de suspensie din ce in ce mai hipoton (prin scaderea
concentratiei de NaCl) si se apreciaza concentratia de NaCl
la care Hb paraseste hematiile si coloreaza
supenatantul in roz = rezistenta osmotica minima
cuprinsa intre 0,46 -0,42 g Na Cl % si concentratia la care
eritroliza este totala =
rezistenta osmotica maxima cu valori de 0,34 - 0,28 g NaCl
%.
In sange, plasma si eritrocitele se gasesc in echilibru
osmotic, adica lichidele separate de membrana eritrocitara sunt
izotonice - au aceeasi presiune osmotica. Scaderea
concentratiei substantelor cristaline dizolvate in plasma
(presiune osmotica mica) va crea intre plasma si eritrocit
un gradient de apa care va determina un flux crescut de H2O
spre hematie, (aceasta intrand prin osmoza). In aceasta
situatie, eritrocitul devine globulos, umflat, membrana nu rezista la
presiunea dezvoltata si se rupe eliberand Hb in plasma (figura
4.7). Introducerea eritrocitului intr-un mediu hiperton va fi urmata de
fuga apei din hematie, care se ratatineaza, adica se sbarceste.
Rezistenta
osmotica este scazuta in majoritatea anemiilor hemolitice.
ANOMALIILE MORFOLOGICE ALE ERITROCITELOR
Variatii de marime
Microcitele
sunt eritrocite mature cu diametrul de 5-6μ, grosimea medie 1,5 μ,
volumul de 60-80 μ3 , suprafata de 100-120 μm2.
VEM, CHEM si HEM sunt scazute (plansa 1).
Fiziologic apar la
copiii in varsta de 2-5 luni, iar la adult sunt normale daca apar in
numar pana la 20% din totalul hematiilor.
Datorita volumului
scazut si/sau incarcarcarii reduse cu Hb au un aspect
de zona clara extinsa central. Cand aceasta zona clara
este extinsa, eritrocitele poarta denumirea de anulocite.
Prezenta
microcitelor pe frotiul MGG este caracteristica anemiilor hipocrome,
feriprive (deficit de fier) dar apar si in talasemii, anemii hemolitice,
etc.
Macrocitele sunt eritrocite mature cu dimensiuni peste
valorile normale. Au diametrul de 8-10μ, volumul peste 100 μ3.
VEM si HEM sunt crescute, CHEM este normal. Apar intens colorate pe frotiu
prin redistribuirea Hb, desi CHEM este normal.
Fiziologic la adult
sunt prezente pe frotiu, pana la 20% din totalul eritrocitelor, iar la nou-nascut in primele 2 saptamani.
Macrocitoza
patologica apare in toate situatiile in care creste numarul
de reticulocite si in anemia din insuficienta hepatica, anemiile
toxice (intoxicatii cu Pb, Au, As), anemiile din neoplasme sau de
cauza renala.
Megalocitele sunt eritrocite mature, de talie foarte
mare. Au diametrul de 12-15 μ, volumul de 120-140 μ3 .VEM
si HEM sunt crescute, iar CHEM este normal. Pe frotiu apar eliptice,
fara zona clara central (desi CHEM este normal),
aparent hipercrome. Curba Price - Jones este deviata la dreapta.
Cauza aparitiei
lor o reprezinta alterarea profunda a mecanismelor de proliferare, ca
urmare a deficitului de vitamina B12 si/sau acid folic).
Megalocitoza este caracteristica anemiei Biermer.
Schizocitele sunt fragmente de eritrocit cu
diametrul de 2-4 μ, de forma rotunda sau neregulata,
hipocrome. Ele sunt fragmente de eritrocite patologice (megalocite,
poikilocite, etc.) sau de eritrocite normale aparute prin fragmentarea
mecanica a acestora. Au o durata de viata scurta.
Sunt caracteristice
pentru anemiile hemolitice traumatice - proteze valvulare, proteze vasculare,
microangiopatii, dar apar si in anemia feripriva, anemii
megaloblastice sau in anemiile induse de chimioterapia anticanceroasa.
Anizocitoza semnifica prezenta pe
acelasi frotiu a hematiilor cu marimi diferite. Pe curba Price -
Jones, anizocitoza se manifesta prin largirea bazei curbei, turtirea
varfului si devierea sa la dreapta sau stanga dupa predominanta
macrocitelor sau microcitelor.
Variatii de forma
Alterarea formei apare
ca urmare a unei patologii membranare, a prezentei unor hemoglobine
patologice sau prin actiunea unor factori interni hematiei.
Eliptocitele /
Ovalocitele sunt
eritrocite de forma ovala, avand doua diametre diferite cu 1-1,5
μ. Prezinta rezistenta osmotica crescuta.
Sunt caracteristice
eliptocitozei ereditare; secundar apar in anemia feripriva, Biermer, neoplazica.
Platicitele /
Leptocitele sunt
hematii cu diametrul longitudinal crescut si grosime redusa. Stroma
are un volum normal, dar Hb este in cantitate redusa. Apar in talasemii,
micro si macrocitoze, etc.
Eritrocitele in "semn
de tras la tinta" sau "palarie de mexican" sau
"saiba" (target-cell / sombrero) prezinta o membrana ondulata
care acopera o cantitate mai mica de Hb la periferie si o
cantitate mai mare in centru.
Apar datorita
gruparii in agregate a moleculelor de Hb si cristalelor de Hb care
dau rigiditate membranei. Se intalnesc in talasemia majora,
hemoglobinopatia C si alte anemii.
Sferocitele sunt hematii cu volum normal, cu
diametrul longitudinal scazut si cu cel transversal crescut (Φ
≤ 6 μm). Aspectul sferic apare prin pierderea de substanta
din membrana.
Se intalnesc in
sferocitoza ereditara (boala Minkovsky - Chauffard), anemii hemolitice
si inainte de hemolize brutale.
Cauza aparitiei
lor este un deficit in structura spectrinei.
Acantocitele/picnocitele/eritrocite
crenelate/echinocite(arici de mare) sunt eritrocite crenelate,
asemanatoare frunzelor de acant. Au pe suprafata lor numeroase
escrescente terminate prin spiculi.
Cauza aparitiei
lor se datoreaza lipsei unei β-lipoproteine din membrana sau
tulburari ale glicolizei. Apar si sub actiunea unor toxine in:
uremie, ciroza, alcoolism cronic, malnutritie severa, etc.
Drepanocitele sunt hematii in forma de
secera care apar datorita polimerizarii unei Hb patologice (Hb
S) sub forma de bastonase helicoidale, care dau rigiditate membranei.
Aceste eritrocite nu pot trece prin capilarele inguste, aparand
obstructii ale acestora (care se manifesta clinic prin infarcte
tisulare) si hemolize. Anemia drepanocitara se mai numeste
si siclemie.
Knitocitele /
Eritrocitele ciupite sunt eritrocite din care s-a desprins un fragment. Au aspect de
coaja de ou sau de semiluna.
Cauza aparitiei o
reprezinta fragilitatea intrinseca, supunerea eritrocitelor unor
traumatisme datorita protezelor valvulare, HTA, microangiopatiilor.
Poikilocitoza reprezinta prezenta pe
acelasi frotiu de eritrocite cu forme negeometrice, neclasificabile sau
variate. Acestea au forma de para, racheta, corn,
picatura, lacrima, virgula, etc.
Poikilocitoza
semnifica o eritropoieza intensa si intotdeauna
patologica. Apare in anemii hemolitice grave, anemii carentiale
severe, eritropoieza extramedulara, mieloscleroza, leucemii,
neoplasme, etc.
VARIATII DE CULOARE
Astfel de variatii
apar in cazul persistentei unor organite: ribozomi (tenta
albastruie), incarcare slaba cu Hb, cresterea grosimii
eritrocitului.
Eritrocite hipercrome - notiune falsa pentru
ca incarcarea E cu Hb nu poate depasi 34%. Acesta este un
aspect specific eritrocitelor din anemia Biermer.
Eritrocitele hipocrome au un aspect mai palid decat
eritrocitele normale. Ele insotesc anemia hipocroma feripriva.
Anulocitele au un
aspect foarte palid si prin concentrarea Hb la periferie capata
aspect de inel
Eritrocitele hipocrome
pot apare si in talasemii, anemii grave (neoplasme, leucemii)
Eritrocitele bazofile provin din eritroblastii care
si-au pierdut nucleul inainte de incarcarea cu Hb. Au o tenta
bazofila, care este data de continutul in ARN. Ele corespund
reticulocitelor.
Eritrocitele
policromatofile
sunt eritrocite care prezinta nuante de la roz-cenusiu la
albastru cenusiu si albastru-violet.
Coloratia este
data de incarcarea variabila cu Hb (coloreaza in roz)
si de prezenta organitelor celulare (coloreaza in albastru).
Semnifica o
regenerare medulara intensa (in anemii) sau o tulburare a
eritrodiabazei (mieloscleroza).
Anizocromia reprezinta prezenta pe
acelasi frotiu a eritrocitelor hipocrome si normocrome (incarcare
inegala cu Hb). Semnifica o tulburare a eritropoiezei.
Incluziile eritrocitare
Incluziile eritrocitare
reprezinta formatiuni cu forma, origine si
semnificatie biologica diferita.
Corpusculii
Howell-Jolly
sunt condensari de cromatina nucleara sau fragmente de cromozomi
ramase in E. Apar ca niste granulatii rotunde, dense,
albastru-violet, unice sau in numar redus (MGG)
Se intalnesc in: anemii
grave, mai ales cele megaloblastice, regenerari eritrocitare intense
si dupa splenectomii.
Granulatiile bazofile
sunt
granulatii mici, albastre sau albastre - violacee, care apar pe toata
suprafata E. (MGG). Contin agregate ribozomale, bogate in ARN,
mitocondrii si fier neheminic.
Apar in
intoxicatii cu metale grele (Pb), talasemii sau alte defecte in sinteza Hb.
Inelele Cabot sunt proteine filiforme ale fusului
mitotic, care au ramas in celula. Apar ca niste formatiuni
filiforme sub forma de linie, circuit inchis (cerc, 8, semiluna,
etc.) de culoare rosie.
Semnifica o
tulburare a mitozei si metabolismului celular (intoxicatii cu Pb,
anemii grave, regenerari intense ale liniei eritrocitare, stadiile
avansate ale neoplasmelor).
Corpii Heinz sunt agregate de Hb degradata
oxidativ. La coloratii supravitale (crezil violet, albastru de Nil, violet
metil, albastru briliant de crezil) apar ca particule de dimensiuni mari,
colturoase.
Apar in:
hemoglobinopatii cu Hb instabila, enzimopatii, dupa splenectomie,
dupa unele medicamente oxidative.
Tactoizii reprezinta cristalele de Hb din
siclemie, care apar prin polimerizarea HbS. Apar ca niste bastonase
helicoidale rigide care la concentratii scazute de O2
devin vascoase si precipita.
Prezenta Hb S
poate fi depistata prin testul de siclizare (eritrocitele suspecte sunt
incubate la presiune partiala scazuta de O2 si in cazul unui rezultat
pozitiv hematiile iau forma de secera)
Siderozomii sunt granulatii de Fe
nehemoglobinic.
Hemoliza
fiziologica.
In cele 120 de zile de viata, eritrocitele
circulante sunt supuse la intense solicitari fizice si chimice, care
le degradeaza treptat 'zestrea' biochimica
initiala si le imbatraneste.
In acest timp, eritrocitele vin si pleaca de la inima de
aprox. 500.000 ori, parcurgand un drum de cca. 150 - 200 km. din care aproape
jumatate este format din teritorii vasculare cu lumen mai mic decat propriul
diametru.
O buna parte din viata lor si-o petrec in vase sanguine
cu structura sinusoidala. Este cazul sinusoidelor splenice (cu un
diametru de 2-3 m)
unde eritrocitele sufera deformari impresionante, care sa le
permita reintrarea in circulatie
(figura 4.6). Traversarea acestor teritorii se face pentru un element
tanar in 30 sec., in schimb celulele imbatranite sau cu defecte pot
fi retinute ore intregi in mediul ostil al circulatiei splenice (pH
acid, hipoxie, hipoglicemie), unde pot fi in cele din urma lizate.
Un adult normal ar avea cca. 25.000 miliarde de eritrocite, dintre care
zilnic se distrug prin eritroliza fiziologica 200 miliarde adica
1/120 din masa totala de eritrocite. Aceasta rata de
distructie zilnica nu duce insa la scaderea numarului
de eritrocite circulante, deoarece in conditii fiziologice intre
erotropoieza si eritroliza exista un echilibru dinamic.
Constatarea ca macrofagele ataca si fagociteaza
numai eritrocite imbatranite si/sau alterate sugereaza ca
in decursul vietii se produc alterari structurale si
functionale care permit recunoasterea de catre macrofage.
Progresele inregistrate de tehnicile biochimice si microscopice au permis
elucidarea multora dintre cauzele senescentei eritrocitare, permitand
evidentierea unor alterari morfologice sau biochimice, care timp de
multi ani au fost doar intuite.
Alterari
morfologice. Eritrocitul
imbatranit este mai putin flexibil, isi pierde forma caracteristica de disc biconcav, manifestand tendinta spre sferocitate (sfera cu
spiculi), urmarea fiind scaderea volumului eritrocitar mediu, pierderea
supletei si a capacitatii de a se deforma, fragilitate
mecanica si osmotica crescuta. Cauza acestor
transformari pare sa fie scaderea cantitatii de ATP,
care duce la alterarea retelei de spectrina si actina,
proteinele din retea formand agregate.
Modificarile
biochimice sunt
consecinta degradarii sistemelor enzimatice, eritrocitul neavand
posibilitatea sa le inlocuiasca prin noi sinteze. Au fost
evidentiate nenumarate alterari enzimatice, cele mai multe
interesand enzimele glicolitice, a caror degradare este urmata de
diminuarea glicolizei totale si a activitatii suntului
hexozomonofosfatilor, precum si diminuarea sintezei de ATP.
Deficitul de NADH si de grupari sulfhidrice va avea efecte
negative asupra sistemelor peroxidazice de detoxifere, fiind favorizata
astfel oxidarea Hb in methemoglobina. Continutul crescut de
methemoglobina (prin fixarea stabila a O2) adanceste
hipoxia la nivelul sinusoidelor, agraveaza perturbarile metabolice
si accelereaza captarea eritrocitelor.
Toate anomaliile descrise duc in final la pierderea
capacitatii eritrocitului de a transporta oxigen. Fara
valoare functionala, eritrocitul devine un element inutil, care va fi
indepartat din circulatie.
Mecanismul exact prin care celulele sistemului monocit macrofag sunt
directionate spre eritrocitele in 'agonie' este incert. Se
stie insa ca eritrocitele senescente sau degradate de
diferiti agenti fizici si chimici manifesta un tactism
intens pentru fagocite.
Sediile hemolizei.
Ertrocitele imbatranite sunt captate si fagocitate atat de
catre macrofagele tisulare si circulante (monocitele), precum si
de catre granulocitele neutrofile si eozinofile (figura 4.8).
Cum macrofagele sunt raspandite in toate tesuturile,
eritrofagocitoza se va desfasura cu intensitati diferite in
orice tesut, existand insa teritorii de electie si anume
zonele de staza sau circulatie incetinita, unde macrofagele se
gasesc in numar foarte mare - sinusoidele medulare, splenice,
hepatice.
Cercetarile experimentale au evidentiat ca la iepuri,
eritroliza are loc predominant in M.O.H. si accesoriu in ficat si
splina, in timp ce la sobolani procesele de eritoliza au
aceeasi intensitate in ficat si splina ca si in M.O.H.
Cunostintele acumulate au permis renuntarea la conceptul
ca splina ar fi 'cimitirul hematiilor imbatranite', aducand
in prim plan importanta M.O.H. si a ficatului. In conditii
normale, rolul splinei este secundar, un argument de necontestat fiind durata
normala de viata a hematiilor in conditiile splenectomiei.
In conditii patologice, aceste raporturi se schimba, splina
devenind sediul principal al eritrolizei.
Indiferent de organul in care se produce, eritroliza se
desfasoara in majoritate (90%)
extravascular si doar intr-un mic procent intravascular (10%).
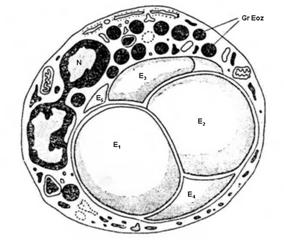
Figura
nr. 4.8. Eozinofil care fagociteaza mai multe eritrocite.
N -
nucleul in desaga;
Gr
Eoz - granulatii eozinofile; E1 - E5 -
eritrocite in diferite faze de digestie (microscopie
electronica, dupa M. Bessis).
|
|
Mecanismul
eritrolizei extravasculare (intratisulare)
A fost elucidat prin tehnici de microscopie electronica,
microscopie in contrast de faza, microcinematografie si curprinde 3
etape.
- etapa de aderenta sau acolare in care celula fagocitanta,
adera la eritrocit;
- etapa de inglobare sau endocitara, in care fagocitul emite
pseudopode care inconjoara eritrocitul ca o palnie sau gura
deschisa, si-1 capteaza in citoplasma(figura 4.9);
- etapa de digestie, in care enzimele din lizozomi vor distruge
membrana eritrocitara cu eliberare de Hb.
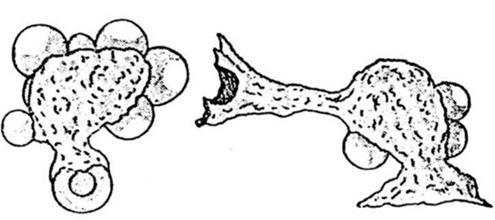
Figura nr. 4.9. Doua macrofage care au
"ingurgitat" mai multe eritrocite, aflate in diferite momente ale digestiei.
Macrofagul din dreapta emite un pseudopod terminat printr-o "palnie" catre
eritrocitul deja captat de celalalt macrofag (dupa o fotografie la
steroscan).
Din catabolismul constituentilor membranari si ai stromei
rezulta: aminoacizi, lipide si fosfolipide care intra in
circuitele metabolice ale organismului.
Degradarea Hb consta in
separarea globinei de nucleul feroprotoporfirinic (hemul) si hidrolizarea
ei pana la aminoacizi, care intra in fondul comun metabolic (figura
4.10). Zilnic din catabolizarea a 6,25g de Hb se elibereaza 6g de
aminoacizi.
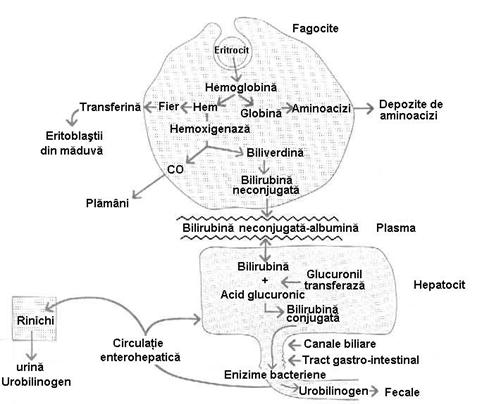
Figura nr. 4.10. Schema hemolizei extravasculare (intratisulare)
Nucleul protoporfirinic este
desfacut sub actiunea unei hemoxigeneze microsomale in: CO,
Fe si biliverdina (coleglobina).
▪ CO se combina cu Hb
formand carbaminhemoglobina si se elimina prin plamani.
▪ Fierul, in macrofag este
depus sub forma de feritina, apoi in sange se leaga de
transferina si este transportat (90%) la M.O.H. unde serveste la
sinteza precursorilor eritrocitari. Restul este depus in depozite sub
forma de feritina.
▪ Biliverdina este redusa
la Br.libera sub actiunea biliverdinreductazei. Br. libera
(indirecta) se leaga de o albumina plasmatica, rezultand
bilirubina prehepatica. In hepatocit: bilirubina prehepatica se
conjuga cu acidul glucuronic, in prezenta enzimei glucuroniltransferaza
si rezulta bilirubina conjugata sau hepatica. Deficitul de
glucuroniltransferaza caracterizeaza sindromul Criggler-Najjar. Br
conjugata ajunge la colecist, apoi prin bila este eliminata in
intestin, unde in prezenta bacteriilor intestinale se transforma in
urobilinogeni (urobilinogen, mezobilinogen, stercobilinogen), care, fie se reabsoarbe si prin circulatia
portala ajunge la ficat, fie se elimina prin fecale, pe care le
coloreaza (stercobilinogen). O parte din urobilinogenul ajuns la ficat
trece in sange si ajunge la rinichi, dupa care este eliminat sub
forma de urobilinogen.
Mecanismul eritrolizei
intravasculare. Cand
liza hematiilor se produce intravascular (in circulatia rapida), Hb
eliberata direct in sange disociaza in dimerii a si b, care sunt legati rapid de o
globulina plasmatica numita haptoglobulina
(Hp). Complexul Hb - Hp, avand GM mare nu poate trece filtrul glomerular
ramanand in sange, de unde este captat de celulele sistemului monocit -
macrofag si catabolizarea Hb se produce pe cai normale (figura 4.11).
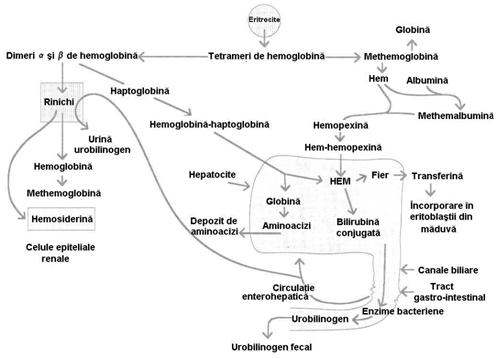
Figura nr. 4.11.
Hemoliza intravasculara. Etapele intermediare ale conversiei hemului in
bilirubina conjugata sunt redate la schema hemolizei extravasculare
In situatia in care hemoliza intravasculara este intensa
si cantitatea de Hb eliberata depaseste capacitatea Hp
de a o lega, sau in conditiile in care exista o scadere
patologica de Hp, dimerii a si b se filtreaza glomerular. Unii dintre ei
sunt captati (reabsorbiti) de celulele tubulare renale in care Fe
este dispus ca hemosiderina, dupa care aceste celule se descuameaza
si se elimina prin urina. Prezenta hemosiderinei in
urina semnifica o cantitate mare de Hb libera circulanta.
Cand capacitatea de reabsorbtie tubulara este
depasita, dimerii se elimina direct prin urina - apare
hemoglobinuria, care semnifica hemoliza intravasculara
severa.
O parte din Hb libera plasmatica este oxidata in metHb
si in aceasta situatie hemul va fi fixat de hemopexina sau albumine. Hemopexina este o glicoproteina
plasmatica cu afinitate pentru gruparile hem de care se leaga
molar. Complexele formate vor fi captate si catabolizate pe cai
normale de sistemul manocit-macrofag, mai ales la nivelul ficatului.
Eritroliza
patologica. Distrugerea precoce a eritrocitului cu defecte membranare
sau de continut, sau chiar a E normale morfo-functional, dar supuse
unor agresiuni extrinseci, este caracteristica anemiilor hemolitice.
Eritolizele patologice sunt clasificate de unii autori in intratisulare si intravasculare. In eritoliza
intravasculara, eritrocitele sunt distruse in circulatie, iar Hb este
eliberata in plasma, in timp ce in cea intratisulara, mai
apropiata ca mecanism de hemoliza fiziologica, eritrocitele sunt
distruse in celulele sistemului manocrit-macrofag.
Cei mai multi autori admit clasificarea hemolizei patologice in intracorpusculara si extracorpusculara, dupa cum,
cauza hemolizei tine de un defect al eritrocitului sau de alterarea
mediului in care traiesc eritrocitele.
Anemiile hemolitice intracorpusculare. Cele mai multe sunt congenitale si se
datoreaza:
I. Unor defecte genetice sau dobandite ale membranei eritrocitare.
II. Unor deficite genetice sau dobandite ale sistemelor enzimatice.
III. Unor defecte cantitative sau calitative ale Hb.
I. Din prima grupa fac parte:
Sterocitoza
ereditara este
boala Minkowski - Chauffard sau microsferocitoza. Este transmisa autosomal
si se manifesta numai in formele heterozigote. S-ar datora unor
defecte in structura proteinelor membranare, in special a spectrinei si
ankirinei. Clinic: anemie, splenomegalie, icter. Pe frotiul sanguin se
constata prezenta microsferocitelor, eritrocite cu f intre 6-7 m. Hb este scazuta, iar VEM, HEM,
CHEM sunt de asemenea scazute. Rezistenta osmotica este si
ea scazuta
Hemoglobinuria
nocturna paroxistica se caracterizeaza prin hemoliza intravasculara
cronica, cu caracter intermitent, datorata unui defect inca
neprecizat, se pare dobandit al membranei, dar care sensibilizeaza
eritrocitul la actiunea distructiva a complementului.
II. Deficite
enzimatice. Teoretic
toate enzimele eritrocitului pot provoca deficite mai mult sau mai putin
severe, detasandu-se insa deficitul enzimelor cu roluri metabolice
importante cum ar fi: enzimele caii glicolitice si ale cailor
derivate din glicoliza (suntul Rapaport-Luebering si suntul
pentozelor), deficite ale enzimelor care intervin in metabolismul
glutationului. Mai frecvent apare deficitul de methemoglobinreductaza,
ereditar sau dobandit, care determina boala numita methemoglobinemie.
III. Anemii hemolitice
prin defecte ale sintezei Hb (vezi capitolul Hemoglobina).
Anemiile hemolitice extracorpusculare
Cele mai multe sunt dobandite
si sunt consecinte ale agresiunii realizate de factorii fizici,
chimici, biologici din plasma. Marea majoritate a acestor factori
actioneaza prin 2 mecanisme:
A Actiune toxica
directa asupra unor componenti membranari sau ai
citosolului.
▪ Metalele grele - Pb produce hemoliza prin blocarea
gruparilor SH ale proteinelor din membrana eritrocitara. In
intoxicatia acuta, hemoliza este rapida si masiva, cu
Br. indirecta crescuta si icter, iar in cea cronica, durata
de viata a eritrocitului este scazuta prin scaderea
rezistentei osmotice si mecanice, deoarece Pb se fixeaza pe S
eritrocitului si provoaca leziuni membranare. Intoxicatia cu
arsenic este insotita si ea de anemie hemolitica, dar
mecanismul hemolitic este incert.
▪ Eterul, benzenul si substantele inrudite (trinitrotoluenul, DDT-clorfenoton),
cloroformul, precum si alcoolul - dizolva lipidele din membrana
si stroma.
▪ Acizii si bazele - patrund in hematie, bazele
degradand proteinele stromale.
▪ Agentii oxidanti - produc hemolize, uneori foarte
grave deoarece prin interactiunea O2 cu Hb rezulta O2-
(ionul superoxid) si H2O2
(peroxidul de O2) in cantitate mare, care
depaseste capacitatea protectoare a glutationului redus,
oxideaza gruparile SH ale globinei si produc precipitarea ei, cu
formarea corpilor Heinz. Oxidarea se exercita asupra hemului, rezultand
metHb, asupra globinei, care precipita, sau asupra lipidelor membranare, a
caror alterare duce la pierderi de membrana. Prin acest mecanism
actioneaza: - cloratii, mai ales de K si Na, care
oxideaza direct Fe din Hb; nitritii - de Na, de amil, nitroglicerina
folositi in industria farmaceutica; pigmentii anilinici -
folositi drept coloranti, in industria alimentara sau
farmaceutica; nitroclorbenzenul, precum si naftalenul si
paradiclorbenzenul, care intra in
constitutia unor insecticide si pot fi ingerate accidental, mai ales
de copii.
▪ Toxine animale sau vegetale: veninul unor serpi - vipera
Ruselli, sarpele cu clopotei -contine fosfolipaza A, care
ataca fosfolipidele membranare, generand lizolecitine si
lizocefaline, substante cu actiune puternic hemolizanta.
Sindromul hemolitic este de obicei de mai mica intensitate decat cel neurologic
si cel hemoragic. Veninul de albine si viespi in cantitate mare
produce hemoliza deoarece contine licitinaza. Veninul unor
paienjeni - vaduva neagra are de asemenea actiune
hemolizanta.
Saponina, digitonina sunt toxine vegetale
hemolizante 'in vitro' prin actiunea asupra lipidelor
membranare, mai ales asupra colesterolului. Anumite ciuperci, ca Helvella
esculenta contin acidul helvelic cu actiune hemolizanta,
intoxicatiile cu aceasta ciuperca fiind uneori mortale.
▪ Medicamente - cu actiune
hemolizanta directa sunt considerate unele sulfamide, mai ales cele
folosite in tratamentul leprei, vitaminele K de sinteza, fenacetina.
▪ Agenti biologici ca bacterii, virusuri,
paraziti pot produce hemoliza, de cele mai multe ori prin
actiune directa, rareori prin mecanism imunologic. Bacilul perfringes
- secreta o hemolizina si o a - lecitinaza
care produce orificii in membrana eritrocitului prin distrugerea
fosfolipidelor. Toxinele secretate de acest bacil produc distrugeri ale proteinelor din membrana eritrocitului. Leziuni
asemanatoare produc si: stafilococul prin hemolizina a, streptococul
prin streptolizina O, pneumococul si colibacilul. Dintre paraziti cel
mai incriminat in patogenia anemiilor hemolitice este Plasmodium malariae,
mecanismul hemolizant fiind inca mcert.
▪ Traume suferite in circulatie: traumatism extern in timpul
activitatii fizice - hemoglobinuria de mars cand E. sunt
distruse prin fragmentare (prevenire: incaltaminte cu talpi
moi); hemoliza cardiaca - apare la 10% dintre pacientii cu proteze
valvulare (prin fragmentare); traume
datorate alterarii peretelui vascular: HTA, proteze vasculare,
microangiopatii.
B. Hemolize datorate
declansarii unor mecanisme imunologice.
Accidentele
hemolitice datorate incompatibilitatii de grup sanguin.
▪ Medicamente: penicilina sau cefalosporinele
declansaza reactii imunoalergice cu formare de atc. (Ig. G)
fata de drogul sensibilizant (eritrocitul fiind doar suportul
reactiei antigen - atc). Antigenul se fixeaza ferm pe o
glicoproteina suport din membrana eritrocitului, reactia cu atc.
fiind urmata de hemoliza.
Alte medicamente imunoalergizante (sulfamide, rimfapicilina,
insulina, chinidina), se leaga de o proteina substrat si stimuleaza sinteza de atc. de
tip IgM, cu care reactioneaza in plasma cu formarea unor
complexe atg. - atc., care se fixeaza lax pe membrana hematiei. Frecvent,
la acest complex se fixeaza si C'. Mai tarziu atc. se desprinde
impreuna cu medicamentul, dar C' ramane pe membrana eritrocitului
si-l distruge. In acest caz, hematiile sunt 'martori
inocenti' ele nefiind purtatoare de atg.
▪ Anemii hemolitice datorate producerii de anticorpi reactivi la
temperatura corpului: - anemia imunohemolitica cu atc. la cald - Ig.G
- anemia imunohemolitica cu atc. la rece - Ig.M

