PROPRIETATILE NEURONULUI
EXCITABILITATEA
Excitabilitatea
este o insusire generala a tuturor celulelor vii, iar in ceea ce
priveste neuronii reprezinta capacitatea de a reactiona in mod
specific la actiunea unor excitanti. Pentru a definii mai concret excitabilitatea
putem spune ca e proprietatea neuronilor de a reactiona la
actiunea unui stimul sau excitant mai ales la nivelul membranei neuronale
producand anumite modificari fizico-chimice atunci cand stimulul sau excitantul indeplineste
anumite conditii. Stimulul sau excitantul la nivelul membranei neuronale
provoaca modificari in ceea ce priveste permeabilitatea ionilor,
modificari care duc la schimbarea repartizarii acestora de o parte
si de alta a membranei neuronale. In termeni fizici, excitabilitatea este
proprietatea neuronilor de a-si modifica rapid potentialul membranal
de repaus si de a genera un potential membranal de repaus si de
a genera un potential membranal de actiune cand asupra membranei
actioneaza un stimul sau un excitant. Pentru a produce o
excitatie stimulul trebuie sa indeplineasca 4 conditii:
stimulul trebuie sa aiba o
anumita intensitate, care a fost numit intensitate-prag sau liminara si care se defineste ca
intensitatea minima capabila sa provoace excitatia la
acsiunea stimulului. Stimulii care au valoare sub prag se numesc stimul subliminari, iar stimulii care
au valoare peste prag se numesc stimuli supraliminari.
In urma unei singure actiuni stimulul subliminar determina o
depolarizare locala care poarta denumirea de potential local
si care are un rol important in realizarea excitabilitatii.
Daca actiunea cu un stimul subliminar se repeta cu
frecventa mare, potentialele locale se sumeaza si pot
genera un potential de actiune.
stimulul
sau excitantul trebuie sa dureze un interval suficient de timp, sa
actioneze intr-un anumit timp numit
timp util.
stimulul
sau excitantul trebuie sa actioneze cu o anumita bruschete.
Daca intensitatea unui stimul electric creste lent de la zero,
intensitatea va depasi valoarea prag a excitantului fara a
provoca o excitatie. Din aceste considerente excitantul a fost definit
variatia brusca a energiei unui factor de mediu intern sau extern
care produce o excitatie. Scaderea excitabilitatii, atunci
cand intensitatea stimulului creste lent, poarta denumirea de acomodare.
stimulul
sau excitantul trebuie sa atinga o anumita densitate pe unitate
de suprafata. In cazul in care stimularea se realizeaza cu un electrod punctiform
vorbim de o stimulare mai mare pe o unitate de suprafata.
Potentialul membranal de repaus
Daca
se plaseaza 2 mictroelectrozi conectati la un osciloscop catodic pe
suprafata membranei unui neuron vom observa pe ecranul osciloscopului
ca acul sau imaginea indica pozitia zero. Daca unul dintre
microelectrozi strapunge membrana si patrunde in interiorul
neuronului, voltmetrul osciloscopului indica o diferenta de
potential de -70 milivolti ceea ce inseamna ca
potentialul din interiorul neuronului este aproximativ 70 mV, (mai
negativ) decat potentialul din exteriorul acestuia. S-a constatat ca
potentialul unui neuron poate sa ajunga la -90 mV. Datorita
repartitiei inegale a ionilor pozitivi si negativi, de o parte
si de alta a membranei neuronale, membrana este polarizata pozitiv pe
partea externa si negativ pe partea interna. Daca ionii de
Na patrund in celula nervoasa, datorita cresterii
permeabilitatii membranei pentru acesti ioni, peste un timp
oarecare ei sunt eliminati din nou in exteriorul celulei cu forta de
catre asa numitele pompe ionice care elimina ionii de Na din
celule si reintroduc ionii de K, ioni de K care au iesit lent si
pasiv din celula. Cercetatorii au constatat ca membrana
neuronala este de aproximativ 100 de ori mai permeabila pentru ionii
de K decat pentru ionii de Na. De aceea ionii de K au o contributie mai
mare la generarea potentialului membranal de repaus, decat ionii de Na.
Potentialul membranal de actiune reprezinta o depolarizare
rapida a membranei urmata de o repolarizare la fel de rapida
care se propaga de-alungul fibrei nervoase. potentialul membranal de
actiune este expresia electrica a ceea ce numim impuls nervos. Succesiunea de impulsuri nervoase, deci de
potentiale de actiune constituie un influx nervos. In desfasurarea uui potential de
actiune se disting mai multe faze.
O faza latenta;
O faza de depolarizare;
O faza de repolarizare rapida;
Un postpotential negativ;
Un post potential pozitiv.
Faza
latenta
dureaza aproximativ 0,1 milisecunde. Ea incepe in momentul actiunii
stimulului si se incheie cand potentialul de membrana scade de
la valoarea de repaus pana la valoarea pragului de excitatie. In
aceasta faza se deschid canalele pentru ionii de Na
voltaj-dependente, crescand permeabilitatea membranei pentru ionii de Na.
Faza de
depolarizare
dureaza aproximativ 0,8 milisecunde. Ea se intinde din momentul cand
potentialul de membrana atinge valoarea prag de excitatie,
adica de -55 mV si momentul in care atinge o valoare apropiata
de ceea ce noi numim potentialul
nerst +35 mV. Potentialul nerst
pentru un anumit ion corespunde cu acea valoare potentialului de
membrana care previne total fenomenul de difuziune neta, prin
membrana in orice sens a acelui ion.
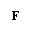 Potentialul nerst se calculeaza
astfel: E = RT log concentratia intracelulara
a ionului / concentratia extracelulara a ionului.
Potentialul nerst se calculeaza
astfel: E = RT log concentratia intracelulara
a ionului / concentratia extracelulara a ionului.
E = potentialul nerst
R = constanta a gazelor
T = temperatura absoluta
F = constanta Faraday
Raportul dintre
concentratia intracelulara si extracelulara pentru ionul de
Na de exemplu este de 10/140= 0,14; log 0,14= -1
RT/F = -61=> E=
61x (-1) = 61
In
aceasta faza ionii de Na navalesc in interiorul celulei.
Fenomenul se numeste depolarizare. Cand valoarea potentialului
membranal de acsiune se apropie de valoarea potentialului nerst
pentru ionul de Na, difuziunea acestuia spre interior este blocata,
datorita respingerii lui de catre sarcinile pozitive de pe fata
interna a membranei. Imediat canalele de Na voltaj-dependente se inchid,
iar permeabilitatea membranei pentru ionii de Na revine la nivel de repaus. Un
canal voltaj-dependent are 2 parti: una activatoare situata la
extremitatea externa a canalului si alta inhibitoare sau
inactivatoare situata la extremitatea interna a membranei. In starea
de repaus a membranei, partea activatoare este inchisa, iar cea
interna este deschisa. Scaderea potentialului de membrana
de la starea de repaus, adica de la +70 mV pana la pragul de
excitatie de -55 mV si in continuare la valor mai mici care pot fi
negative si dupa aceea pozitive duce la deschiderea partii
activatoare si apoi mai lent la inchiderea partii inactivatoare.
Partea inactivatoare se redeschide numai dupa ce potentialul de
membrana atinge in urma repolarizarii valori apropiate de
potentialul membranal de repaus.
Repolarizarea
este rapida
si dureaza aproximativ o milisecunda. Repolarizarea incepe cu
inchiderea de catre partea inactivatoare a canalelor voltaj-dependente
pentru ionii de Na si cu deschiderea canalelor K dependente pentru ionii
de K. Ionii de K difuzeaza din mediul intracelular in cel extracelular in
virtutea gradientului de concentratie. Datorita efluxului crescutd de
ioni de K imediat dupa depolarizarea membranei are loc un inceput de
repolarizare adica potentialul membranal trece de la valoarea
pozitiva la valoarea negativa.
Faza
postpotentialului negativ dureaza 2 pana la 3 milisecunde si se datoreaza
reducerii refluxului de ioni de K prin canalele K voltaj-dependente
datorita inchiderii lor. In aceste conditii repolarizarea membranei
la nivelul potentialului membranal de repaus se realizeaza prin 2
mecanisme:
difuziunea
ionilor de Na din vecinatatea membranei spre interiorul citoplasmei;
prin
activitatea pompei care elimina Na in exteriorul celulei si
reintroduce K in interiorul celulei.
Faza
postpotentialului pozitiv este datorata pompelor ionice, care determina in exteriorul
membranei un exces de sarcini pozitive ceea ce duce sau determina
depasirea potentialului membranal de repaus pana la 10 mV.
In felul acesta se instaleaza o stare de postpotential pozitiv, iar
mai apoi un potential membranal de repaus.
Potentiale locale Prin actiunea unui stimul subliminal asupra unui axon la locul de
actiune se produce o depolarizare usoara a membranei numita
potential local. Cauza acestui potential local este deschiderea unui
numar mic de canale pentru Na voltaj-dependent. Potentialele locale
se propaga pe distante mici, iar amplitudinea si durata
potentialelor locale creste odata cu marirea
intensitatii stimulului subliminal in raport cu potentialul de
actiune care au aceeasi amplitudine si durata indiferent
daca ele sunt provocate de un stimul prag sau supraliminal.
Potentialele de actiune fata de cele locale se supun legii
"totul sau nimic". Potentialele locale nu au o faza refractara,
de aceea efectul stimulului subliminal care actioneaza cu o
frecventa mare se sumeaza. Daca prin suprapunerea
depolarizarii succesive se atinge nivelul critic de -55 de mV, adica
nivelul critic de depolarizare se declanseaza un potential de
actiune. Stimulii de intensitate prag care acctioneaza in faza
refractara a potentialului de actiune nu provoaca un nou
potential de actiune nici chiar prin marirea amplitudinii
potentialului respectiv.
In concluzie, in cazul potentialului de
actiune nu se realizeaza insumarea potentialelor locale.
DETERMINAREA EXCITABILITATII
Pentru a determina excitabilitatea de cele mai
multe ori se utilizeaza stimuli electrici care au o anumita
durata de actiune si o anumita intensitate. Timpul necesar
pentru ca un curent electric sa determine excitatia este cu atat mai
scurt cu cat intensitatea lui este mai mare. Aceasta relatie se
exprima grafic printr-o hiperbola nu numai in cazul tesutului
nervos, ci si in cazul altor celule din organism.
In 1903 savantul francez V. Lapicque a
initiat o metoda de determinare a excitabilitatii ce
implica mai multi parametri. Cei mai importanti sunt: reobaza
si cronaxia.
Reobaza reprezinta intensitatea minima a
curentului electric care declanseaza un potential de
actiune, iar cronaxia este timpul minim
util pentru declansarea unui influx nervos atunci cand se
actioneaza cu un curent electric care are intensitate dubla
fata de reobaza.
t - timp
I - intensitatea
R - reobaza
C - cronaxia
Cronaxia axonilor si a fibrelor musculare pe
care le inerveaza au valori apropiate. In activitatile voluntare
este bine sa nu existe diferente mari intre cronaxia neuronului
si cea a muschiului pe care-l inerveaza. Altfel spus
contractia musculara depinde in mare masura de neuronul
care actioneaza asupra fibrei musculare neuron care trebuie sa
fie intr-o stare de functionare perfecte.
Electrotonusul
In conditii fiziologice potentialele de
actiune sunt generate de diferiti stimuli care pot fi mecanici,
fizici sau chimici. In laboratoarele de neurofiziologie se utilizeaza
stimuli electrici deoarece pot fi cuantificate caracteristicile si se
manipuleaza cu usurinta. In aceste laboratoare, pentru
stimularea electrica, se utilizeaza 2 electrozi, unul negativ si
unul pozitiv. Prin stimularea cu un curent continuu excitatia se produce
doar la inchiderea si deschiderea circuitului electric sau la
variatii bruste ale curentului.
La inchiderea circuitului electric sau odata
cu cresterea brusca a excitatiei se produce o excitatie
inregiunea catodului, iar la deschiderea circuitului sau micsorarea
brusca a excitatiei, aceasta se produce in regiunea anodului. La
inchiderea circuitului la anod pornesc linii de curent. Aceste linii de curent
in majoritatea lor strabatmembrana fibrelor nervoase pana in
interiorul lor unde sunt conduse cu usurinta datorita
electrolitilor si in final ajung la catod.
Liniile de curent de la anod la catod, prin
lichidul interstitial sunt foarte putine datorita acestui strat
de lichid.
(anod) A+← Cl- - anion
(catod) C- ← Na+ - cation
Sub actiunea liniilor de curent sunt
antrenati brusc ionii cei mai mobili din axoplasma. Este vorba de
ionul de K si ionii de Na. Ei ajung de la anod la catod unde se
acumuleaza. Cresterea electropozitivitatii pe fata
interna a membranei in zona catodului inseamna depolarizarea
membranei sau catelectrotonus fapt ce genereaza deschiderea canalelor de
Na voltaj-dependente si generarea unui potential de actiune.
In urma migrarii ionilor de Na in zona
anodului are loc o hiperpolarizare, adica partea interna a membranei
devine mai electronegativa intrucat ionii de Na a plecat de-acolo.
Aceasta hiperpolarizare mai poarta denumirea de anelectrotonus.
Zonele care au un catelectrotonus adica sunt
usor depolarizate au o excitatie mai crescuta, iar cele cu
anelectrotonus sunt usor hiperpolarizate cu o excitatie mai
scazuta fata de normal.
Conductibilitatea
Conductibilitatea este definita drept
proprietatea de a conduce impulsul nervos de la dendrite la corpul neuronului,
iar de aici prin axon la arborezatia terminala a axonului.
La om potentialul de actiune se
propaga mai rapid in fibrele nervoase mielenizate si cu o viteza
mai mica in fibrele nervoase nemielenizate. In fibrele nervoase
nemielenizate viteza de propagare este intre 0,5 pana la 2 m/sec. In
fibrele mielenizate viteza este de 50 de ori mai mare.
In momentul producerii potentialului de
actiune se realizeaza o inversare a polaritatii membranei
pe o lungime intre 16 si 60 de mm. (P. Gherghel, 2001). Astfel in zona
depolarizata membrana devine electropozitiva pe fata
interna datorita patrunderii masive a ionilor de Na, si
electronegativa pe partea externa datorita ionilor de Cl care
raman in cantitate mare in aceasta zona.
Intre zona depolarizata sau activa
si zona polarizata sau inactiva din imediata vecinatate a
zonei excitate apare o diferenta de potential care
determina migrarea sarcinilor pozitive, respectiv a ionilor de Na in
axoplasma din zona depolarizata respectiv activa in zona
polarizata respectiv inactiva.
In lichidul interstitial (la exteriorul
membranei) migrarea se va realiza din zona polarizata in zona
depolarizata. Aceasta migrare a cationilor a fost asemanata
cu anumiti curenti de mica intensitate de catre Robert
Hermann in 1879 si ulterior au luat denumirea de curenti locali
hermann. Acest curenti realizeaza un circuit al carui efect este
diferit pe cele doua fete ale membranei neuronale.
Pe fata interna a membranei sarcinile
pozitive ajunse din zona depolarizata in zona polarizata au efect
depolarizant adica realizeaza catelectrotonus. Ca urmare a
depolarizarii adica a scaderii valorii potentialului
membranal de repaus de la -70 mv la -66 mv se deschid canalele de Na
voltaj-dependente si Na patrunde in axoplasma amplificand
efectul de polarizant al curentilor locali.
Deplasarea cationilor din zona depolarizata
spre cea polarizata intretine efectul depolarizant, amplificator
si astfel depolarizarea membranei in zonele inactive atinge nivelul critic
de -55 mv care este si nivelul prag si se declanseaza un
potential de actiune.
Concluzii:
Potentialele de actiune se propaga
in fibrele amielenice din aproape in aproape cu ajutorul curentilor
locali. Daca pe fata internaa membranei neuronale efectul
curentilor locali este depolarizant si duce la declansarea unui
nou potential de actiune pe fata externa, curentii
locali au o actiune repolarizanta adica sarcinile pozitive se
deplaseaza din zona inactiva in cea activa si
determina restabilirea potentialului membranal de repaus.
Rezulta ca unda de depolarizare a potentialului de actiune
este urmata de o unda de repolarizare.
In fibrele nervoase mielenizate potentialele
de actiune se propaga cu viteza mare in jur de 120 m/sec
datorita particularitatilor morfofunctionale ale acestor
fibre, adica in regiunea intermodulilor axolema este impermeabila
pentru ionii de Na, iar teaca cu mielina are efect izolator impiedicand
iesirea curentilor locali prin membrana in lichidul
interstitial. In schimb in regiunea nodulilor reanvier teaca cu
mielina lipseste si numarul canalelor pentru ionii de Na
voltaj-dependente este de aproximativ 400 de ori mai mare decat membrana fibrelor
amielinice.
Curentii care iau nastere pe fata
interna a membranei neuronale ca urmare a generarii unui
potential de actiune la nivelul unui modul ranvier se deplaseaza
pana la nodulul imediat respectiv, fac un salt si provoaca
depolarizarea membranei generand astfel un potential de actiune.
Curentii locali care iau nastere in urma
depolarizarii membranei pana la o valoare a potentialului de +35
mv pot determina depolarizarea in al doilea modul ranvier care a fost inactiv
daca primul nodul este scos din functiune.
In concluzie pe fata externa a membranei
curentii locali se deplaseaza de la nodulul inactiv la cel activ,
deci prezenta tecii de mielina si particularitatile
legate de densitatea canalelor deNa voltaj-dependente din membrana au permis
marirea distantei la care se deplaseaza curentii locali
si odata cu ei atat depolarizarea cat si cresterea vitezei
de propagare a potentialului de actiune.
DEGENERESCENTA SI REGENERAREA FIBREI NERVOASE
Daca se constata ca un corp al
neuronului a suferit leziuni, moartea lui este inevitabila. Odata
distrusi neuronii, ei nu se mai refac intrucat celula nervoasa nu se
divide. Singura zona a corpului unde celulele nediferentiate dau
nastere la noi neuroni este mucoasa olfactiva si unde neuronii
bipolari daca mor, in timp de 2 luni sunt inlocuiti de 2 neuroni.
Daca fibrele nervoase sunt lezate o parte nu
se mai refac, este vorba de fibrele nervoase din sistemul nervos central, iar
altele din sistemul nervos periferic se refac.
Axonii detasati de corpul neuronilor din
sistemul nervos central mor din cauza unor factori necesari supravieturii
lor care se gasesc numai in corpii neuronilor. Am putea spune ca
axonii nu supravietuiesc detasati de corp asa cum membrele
unui individ nu supravietuiesc detasate de corpul acestuia.
Pentru a sesiza care sunt factorii care
determina moartea fibrelor nervoase izolate sau efectuat mai multe
experiente. De exemplu: s-au lezat cativa axoni din maduva
spinarii a unui soarece de laborator. S-a constatat ca axonii
s-au degradat si si-au pierdut functiile lor. Daca
insa la locul leziunii au fost drefati nervi periferici carora
li s-au extras axonii din teaca Schwann axonii lezati au regenerat in interiorul
tecilor Schwann implantate. In momentul in care insa acesti axoni au
venit in contact cu oligodendrocitele si-au incetat cresterea. In
concluzie celulele Schwann stimuleazacresterea axonilor, iar celulele
gliale respectiv oligodendrocitele o inhiba.
O alta experienta a constat in
realizarea unor leziuni mai ample a maduvei spinarii de la sobolani.
La acestea s-au administrat niste anticorpi specifici unor proteine
care inhiba cresterea axonilor. S-a constatat ca axonii cresc
atata timp cat sunt inhibate aceste proteine care nu permit cresterea
axonilor.
Dupa aceste experiente s-a tras concluzia
ca este posibila regenerarea fibrelor nervoase in sistemul nervos
central. Astazi, in urma unor accidente, prin tehnici de microchirurgie
si cu o medicatie adecvata sunt regenerate partial fibre
din maduva spinarii a omului sau din alte zone ale sistemului nervos
central.
S-a mai constatat ca in urma mortii
fibrelor nervoase in zona respectiva se formeaza niste
substante neurotoxice care nu numai ca afecteaza zona
respectiva, dar si influenteaza activitatea sistemului
nervos. Studiile de neurofiziologie au evidentiat ca exista
totusi o proteina asociata cresterii axonilor care se
gaseste in permanenta in sistemul nervos de la periferia
corpului.
Cresterea fibrelor nervoase este
conditionata insa si de unii factori neurotrofici care
stimuleaza insasi activitatea lor. In 1950 o cercetatoare
Rita Levi Montalcini a descoperit aceasta substanta si
pentru care a primit in 1966 premiul Nobel, substanta care
stimuleaza mai ales cresterea fibrleor nervoase senzitive din
sistemul nervos central.


