Notiuni de genetica moleculara
Genetica
moleculara este acea ramura a
biologiei care studiaza ereditatea organismelor la nivel molecular,
biochimic. La toate organismele materialul genetic este reprezentat de acizii
nucleici (ADN si ARN). Acestia au capacitatea de a inregistra sub o
forma codificata biochimic informatia genetica, care
determina toate caracterele si insusirile organismelor vii.
Structura chimica a acizilor nucleici
Acizii nucleici sunt substante chimice macromoleculare
alcatuite din unitati mai simple denumite nucleotide. O nucleotida este alcatuita dintr-o
baza azotata, un zahar si un radical fosforic. Bazele azotate
din molecula acizilor nucleici sunt de doua tipuri: purinice si
pirimidinice, ele rezultand dintr-un nucleu denumit purina si
respectiv pirimidina. Structura moleculara a ADN-ului a fost
descoperit in 1953 de catre WATSON si colab.
Purina este un tip de baza de azotata,
alcatuita dintr-un heterociclu format din 5 atomi de carbon si 4
de azot, in timp ce pirimidina are o alcatuire ceva mai simpla, fiind
o baza azotata formata dintr-un heterociclu de 6 atomi din care
4 de carbon si 2 de azot. Cele mai importante baze purinice sunt adenina (A) si guanina (G). Acestea sunt prezente
atat in molecula de ADN cat si in cea de ARN. Cele mai importante baza pirimidinice
sunt: citozina (C) si timina (T) in ADN. La ARN, in locul
timinei se afla uracilul (U).
Zaharurile care intra in alcatuirea acizilor nucleici
sunt riboza la ARN si dezoxiriboza la
ADN. Aceste zaharuri sunt pentoze, avand cate 5 atomi de carbon.
Prin unirea unei baze azotate cu un zahar, riboza sau
dezoxiriboza, se obtin asa numitele nucleozide. Prin legarea
nucleozidelor de cate o grupare fosforica se obtin nucleotidele care sunt unitatile de baza ale acizilor
nucleici. Prin inlantuirea nucleotidelor se obtin polinucleotide sau acizi nucleici, in care intra mai mult
de 10 nucleotide.
Acidul dezoxiribonucleic (ADN)
Macromolecula de ADN este bicatenara fiind
formata din doua lanturi polinucleotidice, infasurate
elicoidal in jurul unui ax comun, formand un dublu helix.
Cele doua catene ale helixului de ADN sunt antiparalele
Cele doua lanturi polinucleotidice sunt complementare, in sensul ca intotdeauna o nucleotida care
contine o baza azotata purinica se leaga cu una ce
contine o baza azotata pirimidinica si invers. Ca
urmare in macromolecula de ADN nu exista decat 4 tipuri de legaturi:
A-T, T-A, G-C, C-G.
Structura bicatenara a ADN se realizeaza cu
ajutorul unor punti de hidrogen - duble intre adenina si
timina si triple intre guanina si citozina. Aceste legaturi sunt de natura
electrostatica (fig. x.).
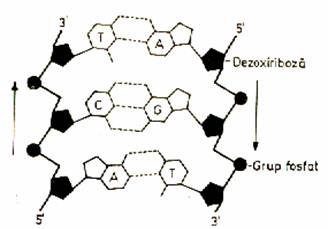
Figura x.Un segment din macro-molecula
de ADN
Replicatia macromoleculei
de ADN
In celulele fiecarei specii se gaseste o
anumita cantitate de ADN, in care se afla sub forma
codificata informatia genetica. Pentru realizarea diviziunii
celulare este necesara, printre altele, dublarea prealabila a
cantitatii de material genetic. In acest fel, celulele fiice vor avea
aceeasi cantitate de ADN cu celula mama.
Sinteza ADN poarta denumirea de replicatie deoarece se realizeaza dupa modelul semiconservativ (fig. x). Prin ruperea
puntilor de hidrogen, moleculele de ADN se separa in cele doua
catene complementare, iar nucleotidele libere din citoplasma se
ataseaza pe baza de complementaritate de catenele vechi. Vor
rezulta doua molecule de ADN bicatenar, fiecare avand o catena veche
(care are rol de matrita) si catena noua
sintetizata.
In acest fel se asigura sinteza noilor molecule de ADN
cu mare fidelitate, moleculele-fiice fiind identice cu moleculele-mama.
Ipoteze privind sinteza ADN considera ca aceasta
s-ar realiza dupa tipul conservativ (sinteza
"de novo") sau dupa tipul dispersiv (dispersarea ADN in
parti componente).
Studiul sintezei ADN in
vivo si in vitro a
demonstrat ca ea se realizeaza dupa tipul semiconservativ propus
de WATSON si CRICK deoarece acest mecanism asigura o inalta
fidelitate in sinteza noilor macromolecule de ADN, identice cu cele vechi.
Tinand seama ca sinteza ADN se realizeaza dupa sistemul
semiconservativ, ca in acest proces cele doua catene servesc drept
matrite pentru catenele nou sintetizate si ca prin acest proces
informatia ereditara este transmisa fidel noilor macromolecule,
procesul de sinteza a ADN a primit denumirea de replicatie
ADN a fost replicat in
vitro, ce catre KORNBERG in 1954, cu ajutorul enzimelor celulare.
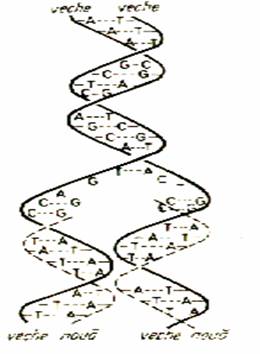
Figura x.
Replicatia ADN dupa tipul semiconservativ
Acizii ribonucleici (ARN)
Acizii ribonucleici (ARN) au, in general, o structura
monocatenara, fiind alcatuiti dintr-un singur lant
polinucleotidic.
Exista mai multe tipuri de ARN, avand functii
diferite:
ARN viral constituie materialul genetic al unor ribovirusuri cum
sunt: virusul mozaicului tutunului (VMT), virusul poliomelitei, virusul gripal
s.a.
ARN mesager (ARN-m) are rol de a copia informatia
genetica a unei catene din macromolecula de ADN si, in felul acesta,
realizeaza ceea ce se cheama fenomenul de transcriptie, o etapa in procesul de decodificare a
informatiei genetice si de sinteza proteica. La ARN-m
succesiunea nucleotidelor este complementara cu aceea a catenei de ADN de
la care a copiat informatia. Greutatea moleculara a ARN-m este
variabila, deoarece si marimea informatiei copiate este variabila.
ARN de transfer (ARN-t) are rolul de a transfera aminoacizii la locul
sintezei proteice. Are o greutate moleculara mica si relativ
constanta, alcatuit dintr-o succesiune de 70-90 de nucleotide. Este monocatenar, cu portiuni bicatenare, care
formeaza o tija si trei bucle mari, fapt care ii confera
forma de trifoi.
ARN ribozomal (ARN-r) intra in alcatuirea ribozomilor
si are rol in sinteza celulara a proteinelor. In molecula sa sunt numeroase plieri neuniforme
datorita legaturilor dintre nucleotidele complementare U-A sau G-C.
ARN nuclear mic (ARN-sn) se gaseste in nucleul eucariotelor,
legat in mod stabil de proteinele nucleare. Acest tip de ARN este alcatuit
dintr-o secventa de circa 100 de nucleotide, avand un rol important
in functionarea nucleului.
Codul genetic si sinteza proteica
In anul 1953, dupa descoperirea structurii
macromoleculei de ADN, s-a lansat supozitia existentei unui cod genetic si ca secventa nucleotidelor de-a lungul
macromoleculei de ADN trebuie sa contina sub forma
codificata informatia genetica a organismelor. Evident ca
s-a pus problema legaturii dintre secventa celor 4 tipuri de
nucleotide, ce contin diferite baze azotate purinice si pirimidinice
(adenina, guanina, citozina si timina) cu secventa aminoacizilor din
catenele polipeptidice.
SAMOV (1954) a emis ipoteza ca in macromolecula
acizilor nucleici se gaseste codificata informatia
genetica necesara sintezei moleculelor de proteine.
Macromoleculele de acizi nucleici contin un numar
mare de nucleotide, astfel ca prin modificarea secventei
nucleotidelor se poate inregistra o enorma cantitate de informatie
genetica. Daca se considera ca o gena este
formata in medie dintr-o secventa de 1000 de nucleotide,
numarul posibil de schimbari in secventa nucleotidelor este
imens, fiind egal cu 41000 sau 10602. Prin inlocuirea
unei singure nucleotide din cele 1000 ale unei gene, se pot produce 3000 de
tipuri de gene alele. Evident ca in felul acesta macromoleculele de acizi
nucleici au o capacitate practic infinita de variatie si
respectiv de inregistrare a informatiei genetice.
Macromoleculele de ADN contin programul sintezei
proteinelor, informatia genetica ce determina ordinea de
succesiune a aminoacizilor. Legaturile dintre secventa nucleotidelor
in ADN si succesiunea aminoacizilor in molecula proteica se
realizeaza cu ajutorul codului genetic. Unitatile de codificare
a informatiei genetice sunt reprezentate de codoni.
Codonul este alcatuit dintr-o secventa de trei
nucleotide din macromolecula de ADN, avand capacitatea de a determina
includerea unui anumit aminoacid in molecula proteica.
Prin codificarea celor 20 de aminoacizi care intra in
alcatuirea proteinelor, exista 64 de codoni, fiecare fiind format
dintr-o secventa de trei nucleotide. Informatia genetica
din moleculele de ADN este mai intai transferata intr-o macromolecula
de ARN mesager (ARN-m), prin fenomenul de transcriptie, dupa care este decodificata si
transformata intr-o secventa de aminoacizi prin translatie (fig. 6.8.)
Intre secventa nucleotidelor din ADN si
secventa aminoacizilor din molecula proteica exista o stransa
corelatie, fenomen denumit colinearitate
Prin combinarea variata a celor 20 de aminoacizi, prin
modificarea secventei lor in catenele polipeptidice, se poate realiza un
numar imens de proteine.
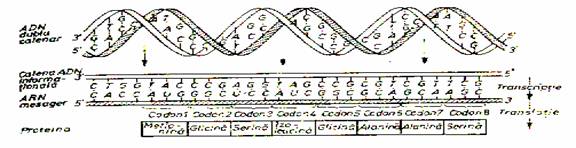
Figura x. Transcriptia si translatia
macromolecului de ADN
Macromoleculele de ADN sunt alcatuite dintr-un
numar mare de nucleotide, care sunt insa de numai 4 tipuri. Prin
modificarea secventei celor 4 tipuri de nucleotide, se poate inregistra,
cu ajutorul codului genetic, o cantitate imensa de informatie genetica.
ADN-ul este deci alcatuit dintr-o secventa de codoni, ce
determina succesiunea aminoacizilor in moleculele proteinelor.
Codul genetic si
caracteristicile sale
Codul genetic este alcatuit din 64 de codoni, cifra
reprezentand totalitatea combinatiilor posibile a celor 4 tipuri de
nucleotide luate cate 3, adica 43. S-a reusit descifrarea
completa a codului genetic, in sensul ca sunt cunoscute toate
tripletele de nucleotide (codoni) din ARN-m ce codifica diferitii
aminoacizi. Acesti codoni sunt formati din 4 tipuri de nucleotide
continand bazele azotate adenina, uracil, guanina si
citozina.
Adevaratul cod genetic continut in macromolecula
de ADN poate fi foarte usor cunoscut prin inlocuirea nucleotidelor din
ARN-m (fig. x.) cu componentele lor: citozina
(C) prin guanina (G), guanina (G) prin citozina (C), adenina (A) prin
timina (T) si uracilul (U) prin adenina (A).
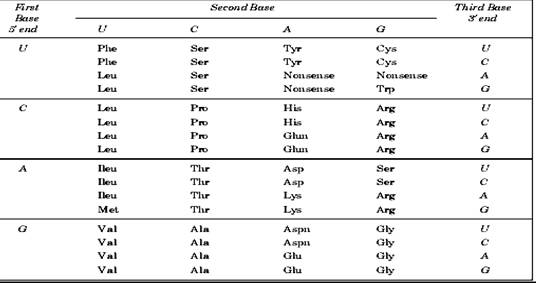
Figura x. Codul genetic ARNm (Phe -
fenil alanina; Leu - leucina; Met - metionina; Val -
valina; Ser - serina; Pro - prolina; Thr - tronina; Ala -
alainina; Tyr - tirozina; Nonsens - STOP; His -histidina; Glum
-glutamina; Asp -acidul aspartic; Lys - lizina; Aspn - asparagina; Cys - cisteina;
Trp - triptofan; Arg - arginina; Ser - serina; Gly -
glicina)
Tinand seama ca exista mai multi codoni
decat aminoacizi (20), s-a dovedit experimental ca mai multi codoni
pot codifica un acelasi aminoacid. Deci codul genetic este degenerat. Datorita fenomenului de degenerare a codului
genetic, un anumit aminoacid poate fi codificat de mai multi codoni
diferiti. Astfel, leucina este codificata de codonii UUA, UUG, CUU,
CUC si CUG.
Codul genetic este nesuprapus, adica
doi codoni succesivi nu au nici o nucleotida comuna si este
fara virgula
(citirea informatiei genetice se
face continuu: intre doi codoni succesivi nu exista semne de
punctuatie). Din totalul de 64 de
codoni ai codului genetic, un numar de 61 codifica cei 20 de
aminoacizi, iar 3 codoni (UAA, UAG si UGA) nu codifica aminoacizii,
ci numai marcheaza sfarsitul unui mesaj genetic (STOP).
Codul genetic este universal. Acest fapt
a fost dovedit prin studiul proteinelor biosintetizate in sisteme celulare
libere, provenite de la bacterii si de la mamifere, sub influenta
unor ARN sintetizati artificial. S-a constatat ca indiferent de
originea sistemului celular liber se obtin aceleasi proteine, fapt ce
constituie argumentul ca aminoacizii ce intra in alcatuirea
proteinelor respective sunt codificate de aceeasi codoni. Tinandu-se
seama de universalitatea codului genetic, s-a dedus ca are o origine
foarte veche, luand nastere in insasi procesul de aparitie
a vietii pe pamant.
Recent, s-a demonstrat existenta unor exceptii de
la "universalitatea" codului genetic. Astfel, in genomul mitocondriilor
(organite celulare de la eucariote) codul genetic prezinta mici deosebiri
fata de cel "universal". De pilda, codonul UGA, care in nucleu
are semnificatia de STOP, in genomul mitocondrial (ADN-mt) codifica
aminoacidul triptofan. Existenta unor deosebiri minore in ADN-mt nu
afecteaza in ansamblu, universalitatea codului genetic, acelasi la
procariotele acelulare si celulare, precum si la eucariote.
Codul genetic este ambiguu, in
sensul ca un anumit codon poate sa includa mai multe tipuri de aminoacizi
intr-o proteina in functie de pozitia lui in catena. Altfel spus,
ambiguitatea codului genetic este caracteristica prin care anticodonul din ARNt
recunoaște doi codoni diferiti din ARNm. De exemplu, codonul GUG aflat la
inceputul mesajului genetic (in molecula de ARNm) este recunoscut de ARNt
pentru formil metionina, pe cand același codon aflat in interiorul
moleculei de ARNm este recunoscut de ARNt pentru valina.
O alta caracteristica a codului genetic este
modalitatea in care se realizeaza descifrarea mesajului genetic
continut intr-o succesiune de nucleotide. S-a demonstrat ca citirea mesajului
genetic se face intr-un singur sens, astfel ca, absenta unei singure
nucleotide (deletia) sau adaugarea unei nucleotide (aditia) face
ca mesajul sa fie citit eronat, in continuare.
Functiile materialului
genetic (transcriptia si translatia informatiei genetice)
Rezultatele cercetarilor privind functiile
materialului genetic pot fi sintetizate in relatia ADN 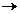 ARN proteine.
ARN proteine.
Conform acesteia informatia genetica se reproduce
prin replicatie si este decodificata (transformata intr-o
proteina sau enzima specifica) prin transcriptie si translatie
Transcriptia constituie fenomenul prin care informatia unei
catene de ADN este transmisa la ARN-mesager (ARN-m), precum si altor
tipuri de ARN.
Translatia inseamna transformarea unei secvente de
nucleotide si respectiv de codoni din ARN-m intr-o secventa de
aminoacizi in catena polipeptidica.
Dupa cum se stie, proteinele, care au un rol
extrem de important structural si functional, sunt foarte variate in
natura. Ele reprezinta, de
regula, circa 50% din substanta uscata a celulelor. Unele
proteine, precum colagenul, au rol in realizarea structurilor organismului, iar
altele, cum sunt enzimele, au rol de a cataliza reactiile metabolice.
Sinteza proteinelor in
vivo se realizeaza pe baza informatiei genetice din acizii
nucleici.
Pana in prezent au fost identificate peste 100.000
proteine diferite, vegetale si animale. In linii mari, toate aceste
proteine sunt alcatuite din variatia secventei a 20 de
aminoacizi.
In procesul de sinteza proteica la nivel celular,
intervin trei tipuri de acizi ribonucleici: ARN-mesager (ARN-m), ARN-ribozomal
(ARN-r) si ARN-solubil (ARN-s), fiecare avand un rol bine determinat.
Rolul ARN in decodificarea informatiei genetice a fost pus in
evidenta de BRACHET si colab. (1950-1955), care au demonstrat
ca in celula sinteza proteica este insotita de o
marire considerabila a cantitatii de ARN.
Recent s-a descoperit ca unele ribovirusuri, la care
materialul genetic este o molecula de ARN, sunt capabile sa determine
in celula respectiva sinteza unui ADN cu ajutorul caruia se
replica. Este pentru prima oara cand se demonstreaza
experimental ca ARN-viral (ARN-v) poate servi ca matrita pentru sinteza ADN-ului. Fenomenul poarta denumirea
de reverstranscriptie sau inverstranscriptie
Etapele sintezei proteice
Dependenta sintezei proteice de informatia
genetica din nucleu a fost pusa in evidenta la alga
monocelulara Acetabularia
mediteraneea. Daca se indeparteaza nucleul celular, biosinteza
proteinelor continua un timp in celula anucleata a algei, insa
cu viteza redusa. Dupa aproximativ 20 de zile, activitatea
biosintetica inceteaza complet si alga anucleata moare.
Daca insa in acest timp se introduce un nucleu in citoplasma
respectiva, in cateva minute alga isi reia pe deplin activitatea.
Prima etapa in procesul de sinteza proteica o constituie
transcriptia informatiei din ADN in ARN-m.
Fenomenul de transcriptie a informatiei genetice
de la ADN la ARN se realizeaza cu ajutorul enzimei ARN polimeraza. ARN-m copiaza informatia genetica numai
a unei catene din macromolecula de ADN.
In celulele procariotelor, ARN-m copiaza
informatia genetica a mai multor gene adiacente care alcatuiesc
un operon. Ca urmare, se sintetizeaza mai multe proteine, de
care celula are nevoie la un moment dat. Pe masura ce se
sintetizeaza moleculele de ARN-m, incepe sinteza catenelor polipeptidice.
La eucariote, ARN-m copiaza, de regula,
informatia genetica a unei singure gene. Acest ARN-m, dupa ce
sufera unele modificari prin eliminarea secventelor
non-informationale, migreaza in citoplasma, unde are loc sinteza
proteica.
Prin transcriptia informatiei genetice se
intelege nu numai sinteza ARN-m ci si a celorlalte doua tipuri
de acizi ribonucleici (ARN-r si ARN-t), care sunt necesari pentru
realizarea sintezei proteice.
A doua etapa a sintezei proteice este reprezentata de translatie, in urma careia o secventa de nucleotide
din ARN-m este transformata intr-o secventa de aminoacizi in
molecula proteica. ARN-m se cupleaza cu ribozomii din citoplasma
formand poliribozomi. Concomitent are loc activarea aminoacizilor (AA) din
citoplasma prin legarea lor de ATP (adenozintrifosfat),
substanta chimica ce serveste ca donator de energie.
Cele trei faze ale biosintezei proteice pot fi redate
sintetic astfel:
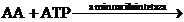 AA~AMP+P~P
AA~AMP+P~P
In aceasta faza un aminoacid oarecare AA este
activat in urma reactiei cu molecula de ATP donatoare de energie sub
influenta enzimelor denumite aminoacilsintetaze. Ca urmare, aminoacidul se
leaga de AMP (adezinmonofosfat), iar doua grupuri fosfat sunt puse in
libertate.
AA~AMP+ARN-t
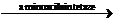 AA~ARN-t+AMP
AA~ARN-t+AMP
In aceasta faza are loc transferul
aminoacizilor activi la ARN-t, sub influenta acelorasi enzime din
etapa precedenta. Cu ajutorul moleculelor de ARN-t aminoacizii sunt
transferati la locul sintezei proteice in ribozomi.
AA1~ARN-t1+AA2~ARN-t2
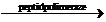 AA1~AA2+ARNt1+ARN-t2
AA1~AA2+ARNt1+ARN-t2
In aceasta faza, aminoacizii, de exemplu,
AA1 si AA2 se unesc intre ei prin legaturi peptidice
cu ajutorul enzimelor peptidpolimeraze. Se
formeaza in acest fel catene polipeptidice, iar moleculele de ARN-t sunt
puse in libertate si sunt reciclate, adica refolosite in procesul
sintezei proteice. De asemenea si ribozomii sunt reciclati in cursul
sintezei proteice.
Gena.
Structura si functii
In conceptia clasica, gena era definita de
catre JOHANNSEN (1909), ca unitate a materialului genetic, localizata
in cromozomi.
In organismele haploide genele se prezinta sub
forma simpla, iar in cele diploide sub forma de alele. Tipul
primar al genei (tipul salbatic) se modifica prin mutatii
formand una sau mai multe alele ce afecteaza acelasi caracter. In
cazul organismelor diploide descendentii primesc numai cate un membru al
perechii respective de alele, de la fiecare genitor.
In conceptia clasica, gena are trei
trasaturi de baza:
uniformitatea
functionala, in sensul ca determina producerea unui efect
fenotipic ;
unitate
mutationala, prin care gena normala ce determina tipul
salbatic se transforma prin mutatii in alela, sau alelele sale,
astfel ca este afectat caracterul respectiv ;
unitate de
recombinare genetica, prin care genele se pot transfera de pe un cromozom
pe perechea sa, prin fenomenul de crossing-over.
Genele, pe baza localizarii lor pot sa fie autozomale si heterozomale. Determinarea pozitiei
unei gene intr-un anumit cromozom a facut posibila alcatuirea hartilor genetice
Sub aspectul manifestarii lor genele pot sa fie
recesive, dominante, codominante, epistatice, hipostatice, complementare sau
indiferente.
In timp ce genele nucleare se transmit
la descendenti si segrega dupa tipul mendelian, genele extranucleare prezinta un tip de mostenire nemendelian.
Intre cele doua tipuri de gene de la organismele eucariote poate exista un
fenomen de interactiune, in sensul ca cele nucleare pot determina
functionarea sau nefunctionarea celor extranucleare, impreuna
putand produce un anumit fenotip.
Apogeul in conceptia clasica despre gena a
fost reprezentata de ipoteza o gena - o enzima,
elaborata de BEADLE si TATUM (1941). Aceasta ipoteza poate
fi sintetizata astfel:
Toate procesele biochimice din organisme sunt controlate
genetic, fiecare reactie dintr-un lant metabolic fiind
determinata primar de o gena. Intre gene si enzimele respective
exista raportul de 1:1. Prin mutatia unei singure gene are loc
blocarea sintezei enzimei corespunzatoare si a reactiei
biochimice catalizata de ea. Ca urmare are loc blocarea intregului
lant metabolic. Ipoteza considera ca fiecare gena
controleaza sinteza, functionarea si specificitatea unei anumite
enzime.
Notiunea de gena a evoluat foarte mult odata
cu aparitia geneticii moleculare si a perfectionarii
metodelor de investigatie la nivel molecular.
Gena in conceptia
actuala poate fi definita drept
un segment al macromoleculei de ADN (sau ARN in cazul unor virusuri), format
dintr-o anumita secventa de nucleotide, care
actioneaza ca o unitate functionala si care
contine secventa nucleotidelor intr-o molecula de ARN-m (transcriptia
genetica) si respectiv a aminoacizilor intr-o catena
polipeptidica (translatia genetica)
Gena este alcatuita dintr-un mare numar de
subloci potential mutabili, dispusi linear intr-o anumita ordine
si intre care poate avea loc fenomenul recombinarii genetice.
Dimensiunea genelor este variabila, in functie de
cantitatea de informatie genetica pe care o poseda. La bacterii
s-a calculat ca in medie o gena este formata dintr-o
secventa de 900-1.500 nucleotide, fiecare aminoacid fiind controlat
genetic de o tripleta de nucleotide denumita codon
Daca o proteina este alcatuita dintr-o
singura catena polipeptidica sau din mai multe dar identice, ea
este codificata de o singura gena. In cazul cand molecula proteica este formata din
mai multe catene polipeptidice nonidentice, ea este codificata de mai
multe gene. De exemplu, hemoglobina A este alcatuita din 4 catene
polipeptidice, identice doua cate doua ( si ), ea fiind
codificata de doua gene diferite.
Sub aspect functional, genele pot fi clasificate in 4
categorii :
1) Gene structurale care determina secventa aminoacizilor intr-o
catena polipeptidica, astfel ca intre secventa
nucleotidelor in acidul nucleic si cea a aminoacizilor din proteina
corespunzatoare se observa fenomenul colinearitatii
2) Gene reglatoare, au rolul de a controla sinteza proteica la nivelul
celulei, in functie de conditiile de mediu extracelular sau de
necesitatile functionale ale celulei.
3) Gene arhitecturale care asigura integrarea proteinelor sintetizate in
structurile celulare.
4) Gene temporare care activeaza cele 3 tipuri de gene anterioare,
pe baza unui program in timp si spatiu, astfel ca in organism se
realizeaza fenomenul citodiferentierii
WADDINGTON (1965), a denumit intreaga serie de procese
biochimice care duc de la gena la caracterul fenotipic respectiv sistem de actiune a genelor
In timp ce la procariote genele sunt formate dintr-o
secventa de nucleotide continua, ce codifica o
secventa de aminoacizi, la eucariote genele sunt alcatuite din
segmente informationale, separate de segmente non-informationale. De
exemplu, gena ce determina sinteza ovalbuminei la gaina este
constituita din "bucati" prezentand intreruperi de secvente
de nucleotide care este transcrisa in ARN-m. Aceasta inseamna ca
in sapte regiuni ale genei exista insertii de secvente de
nucleotide care nu sunt transcrise in ARN-m. Ca urmare, gena este
alcatuita din segmente informationale, separate prin segmente de
ADN non-informational, sau silentios. Secventele de
nucleotide non-informationale incluse in gena respectiva se numesc introni. Au o marime variabila intre 300 si
1.400 perechi de nucleotide. Regiunile intercalare se numesc exoni
si ele constituie cu adevarat gena.
In genomul eucariotelor exista un numar relativ
mic de gene, comparativ cu cantitatea totala de ADN. De exemplu, la om,
s-a calculat ca numarul posibil de gene ar fi egal cu 3 106. In realitate,
numarul de gene este mult mai mic, fiind egal cu 5 104. Aceasta
inseamna ca numai o mica parte din genomul uman este constituit
din gene functionale, in timp ce cea mai mare parte din genom este
constituita din ADN non-informational, cu alte roluri.
Functiile genelor si
dogma centrala a geneticii
Genele au doua functii: autocatalitica
si heterocatalitica. Dogma centrala a geneticii consta in
faptul ca, informatia genetica se gaseste in ADN, ea
putand fi replicata (functie autocatalitica) si
decodificata prin transcriptie si translatie (functie
heterocatalitica). Ca urmare, fluxul de informatii este
unidirectional : ADNARN - proteine.
Functia
autocatalitica a genelor.
Consta in capacitatea lor de a se autoreplica cu mare fidelitate, astfel
ca, in urma procesului diviziunii, celulele fiice mostenesc, de
regula, aceleasi gene, existente si in celula mama.
La organismele pluricelulare, fiecare celula a
organismului poseda totalitatea genelor caracteristice speciei respective,
numai ca in celule specializate functioneaza anumite gene,
restul gasindu-se in stare represata.
La plante s-a demonstrat ca atat celulele diploide ale
organismului, cat si cele haploide, sunt capabile sa regenereze in
culturi celulare intregul organism.
Deoarece la majoritatea organismelor materialul genetic
este reprezentat de ADN si numai in cazul unor virusuri este reprezentat
de ARN, functia autocatalitica a genelor este reprezentata de
procesul de replicatie a acizilor nucleici. Acest proces se
realizeaza dupa tipul semiconservativ, prin care
moleculele de ADN nou sintetizate sunt formate dintr-o catena
polinucleotidica veche si una nou sintetizata. Existenta
complementaritatii nucleotidelor de-a lungul macromoleculei de acizi
nucleici, permite sinteza materialului genetic, cu o mare fidelitate. Numai
asa se poate explica stabilitatea relativa a organismelor vii,
transmiterea informatiei genetice in cursul generatiilor succesive.
Functia
heterocatalitica a genelor.
Consta in capacitatea lor de a determina sinteze specifice de proteine,
enzime si alte biomolecule. Aceasta inseamna ca informatia
genetica pe care o contine fiecare gena este la un moment dat
decodificata si transformata intr-o secventa de
aminoacizi, adica in catene polipeptidice.
In fiecare celula de tip procariot sau eucariot,
genele se gasesc, de regula, intr-un singur exemplar sau sub forma
de alele, in timp ce numarul moleculelor proteice este foarte mare, de
zeci sau sute de mii. Aceasta inseamna ca decodificarea
informatiei genetice continuta in moleculele de ADN se
realizeaza cu o viteza foarte mare, incat celulele pot sintetiza
rapid cantitati mari de molecule proteice. Pentru aceasta exista
un sistem de amplificare, prin care genele sunt copiate in numeroase exemplare
de ARN-m prin fenomenul de
transcriptie si apoi prin translatie sunt transformate in
secvente de aminoacizi, adica in catene polipeptidice.
Recent s-a descoperit ca la eucariote genele cuprind
secvente de nucleotide informationale denumite exoni si secvente non-informationale denumite introni. Astfel, gena este alcatuita din mai multe
bucati, din care numai exonii sunt transcrisi in ARN-m si
informatia lor genetica este decodificata si
transformata in secvențe de aminoacizi. In procesul de
transcriptie intronii sunt eliminati.
De exemplu, gena ovalbuminei, a proteinei din albusul
oualor de gaina, este format dintr-o secventa de 8
exoni si 7 introni, care alterneaza. Ca urmare, genele de la
eucariote sunt mult mai mari decat cele de la procariote, deoarece au secvențe
de ADN non-informational. Acest ultim tip de ADN are rol in procesul de
evolutie prin duplicarea genelor.
Genele de la eucariote au o structura discontinua
sau in mozaic.
Gene suprapuse. La bacteriofagul phix 174, care are un genom circular,
s-au descoperit gene suprapuse. Gena A a acestui virus bacterian este mai mare,
iar gena B de dimensiuni mai reduse, fiind inclus in gene A. De asemenea genele
D si E sunt suprapuse (fig. 6.11.). Aceasta inseamna ca
sinteza ARN-m poate incepe din doua puncte diferite ale aceleasi
gene, astfel
ca
se pot sintetiza doua catene polipeptidice de marimi diferite.
Pseudogene. In genomul unor eucariote au fost evidentiate
pseudogene, adica niste gene relicte, care si-au pierdut
functiile si care au numai rol in mentinerea arhitecturii
moleculare a cromozomilor. Astfel de gene se regasesc in familia genelor
hemoglobinelor, a imunoglobinelor etc. cu care se inrudesc structural. Fiind
nefunctionale, pseudogenele acumuleaza mutatii cu o
frecventa marita.
Elemente genetice transpozabile
Elementele genetice
transpozabile sau transpozonii sunt segmente
de ADN de 750-40.000 perechi de baze, capabile sa circule in cadrul
genomului atat la procariote, cat si la eucariote si sa se
insere in diverse locusuri. Ele pot determina mutatii genice,
restructurari cromozomale, activarea unor gene silentioase,
inactivarea unor gene etc. Traspozonii sunt de doua tipuri :
traspozoni simpli, mai mici
ca dimensiuni si care contin numai gena traspozazei, enzima cu
ajutorul careia se realizeaza transpozitia ;
transpozoni complecsi, de
dimensiuni mai mari, care pe langa gena traspozazei contin una sau
mai multe gene care sunt transpozate dintr-un locus in altul al genomului.
Inducerea transpozitiei se realizeaza in conditii
de stress provocat de agentii fizici (radiatii, socuri de
temperatura) si chimici (substante mutagene, poluante etc.) sau
prin cultura de celule si tesuturi pe medii artificiale. In aceste
conditii se produce o destabilizare a genomului, fapt ce determina o
mare variabilitate genetica. Aceasta permite o capacitate marita
de adaptare a organismelor la conditiile nefavorabile de mediu si
bineinteles supraviatuirea. Elementele genetice transpozabile au un
rol important in evolutia rapida a organismelor vii in conditii
de stress.

