Legile mendeliene ale ereditatii
Atracția oamenilor catre
cunoașterea modului de transmitere a caracterelor ereditare este la fel de
veche ca și umanitatea insași, asemanarea
descendenților cu ascendenții fiind consemnata din cele mai
vechi timpuri. In jurul anului 300 I.C., marele Aristotel a observat ca
anumite particularitați ale parului, unghiilor și chiar ale
mersului, precum și unele caractere inaparente, reapar la
descendenți.
De asemenea, pornind de la constatarea
ca descendența manifesta diferite "diluții" ale
caracterelor genitorilor, intr-o opinie straveche, ereditatea a fost
considerata un proces de mixare, pe baza caruia puteau fi explicate
diferențele și asemanarile dintre indivizi. Inca din
anul 400 I.C., in Grecia, a fost sugerat rolul mediului inconjurator in
producerea variației, idee similara cu cea promovata de "Teoria
caracterelor dobandite", propusa de Lamarck, 22 de secole mai tarziu.
Primele informații care au
facut referire la materialul de baza al ereditații au fost
obținute de catre fermieri, care foloseau diferite tehnici de
hibridare și selecție pentru ameliorarea plantelor cultivate. La
mijlocul secolului al XVIII-lea, taxonomistul suedez Carolus Linnaeus
(1707-1778) și amelioratorii germani Josef Gottlieb Kölreuter (1733-1806)
și Karl Friedrich Von Gaertner (1722-1850) au folosit polenizarea artificiala
la plante, obținand hibrizi in descendența. In 1760, Kölreuter a
publicat o carte in care erau descrise 500 de experimente de hibridare la
plante. Deși observațiile sale erau similare cu cele ale lui Mendel,
interpretarea lor nu era corecta. Gaertner a efectuat experimente similare
la mazare și a obținut raportul de segregare a caracterelor de
3:1, identic cu cel calculat de Mendel, cateva decenii mai tarziu.
La inceputul secolului al XIX-lea, trei
botaniști britanici Knight, Seton și Goss au studiat modul de transmitere
a culorii semințelor la mazare, remarcand ca prin hibridarea
dintre plante cu semințe galbene și plante cu semințe verzi, in
prima generație toate plantele din prima generație au semințe
galbene, iar prin autopolenizarea acestora a fost obținuta
generația a doua formata atat din plante cu boabe galbene, cat
și din plante cu boabe verzi. Prin autopolenizarea plantelor cu
semințe verzi s-a obținut in descendența numai plante cu
semințe verzi.
Experimentele tuturor amelioratorilor
au fost studiate și analizate in detaliu de catre Mendel.
Trebuie subliniat faptul ca, cele
mai importante concepte genetice au fost decelate in urma experimentelor
clasice realizate la plante. La animale, eforturile cercetatorilor au fost
concentrate pe ințelegerea modului de reproducere sexuata. In 1875,
Spallanzani a obținut descendenți, folosind inseminarea
artificiala la caini. Studiind ariciul de mare, in 1875, Oscar Hertwig a
observat ca fecundarea implica unirea nucleului spermatozoidului
și cel al ovulului.
Gregor Johann Mendel s-a nascut
in 1822, la ferma familiei din satul ceh Heizendorf (pe atunci, parte a
Austriei), a carei ocupație principala era crearea de noi
varietați de mar. La un an dupa terminarea colegiului, a lucrat
ca asistent de predare in invațamant, iar din 1848 a devenit
calugar. In 1851, a intrat la Universitatea din Viena pentru a studia
fizica, matematica și științele naturale. Aici a fost
invațat și influențat de Franz Unger (care preda fiziologia
plantelor, cursurile conținand informații semnificative despre
experimentele de hibridare ale lui Kölreuter Gaertner) și Christian
Doppler (cel care a descoperit efectul Doppler). In 1854, dupa terminarea
studiilor s-a intors la Brunn (Brno), in Moravia (Cehoslovacia) unde a
continuat sa lucreze ca preot și ca profesor la școala
superioara.
G. Mendel a inceput cunoscutele sale
experiente de hibridare la plante in 1856
si spre deosebire de predecesorii sai, a colectat și
analizat datele intr-un mod sistematic, matematic.
In primul rand, el a analizat
transmiterea in descendența a cate unui singur caracter, in al doilea
rand a ținut cont de pedigree-ul fiecarei plante, in al treilea rand
a innumarat diferitele tipuri de plante obținute dupa fiecare
experiment de hibridare și in al patrulea rand, a interpretat datele
matematic. Astfel, Mendel a reușit ceea ce predecesorii sai nu au
izbutit.
Mendel a
inceput experientele de hibridare la diferite specii de plante ca: Pisum, Hieracium, Phaseolus, Cirsium,
Linaria, Mirabilis, Zea etc., insa in mod deosebit a facut
cercetari la mazare (Pisum
sativum). Astfel, succesul lui Mendel este atribuit, in parte, și
alegerii materialului vegetal adecvat, mazarea oferind avantajul unei
cultivari ce nu necesita condiții speciale, este autogama,
este potrivita pentru polenizarea artificiala, hibridarea fiind
ușor de realizat și, totodata prezinta perechi de caractere
ce pot fi urmarite cu precizie in generații succesive. De exemplu,
semințele netede și zbarcite sunt forme contrastante ale caracterului
aspectul boabelor. Mendel a studiat transmiterea ereditara a șapte
perechi de caractere, ceea ce inseamna ca a lucrat cu 14 soiuri.
Monohibridismul si legea puritatii gametilor
Monohibridarea consta in hibridarea a doi
indivizi, care se deosebesc printr-o singura pereche de caractere.
Mendel a
realizat hibridari intre soiuri de mazare care prezentau caractere
morfologice diferite, precum culoarea cojii semințelor (colorata sau
alba), consistența pastailor (tare sau moale), culoarea
pastailor (verde sau galbena), aspectul semințelor (netede
sau zbarcite), culoarea cotiledoanelor (galbena sau verde), poziția
florilor (axilara sau terminala) și inalțimea
plantelor (inalte sau pitice).
Primele
experimente ale lui Mendel au explicat cum segrega o gena in
descendența.
Prin
hibridarea dintre o planta de talie inalta (genitorul matern),
homozigota, cu o planta de talie pitica (genitorul patern),
homozigota, a obținut in prima generație (F1)
exclusiv plante inalte, și a denumit cu termenul dominant caracterul
planta inalta, care a dominat in prima generație și cu
termenul recesiv, caracterul planta pitica, care a
ramas ascuns (latent) in generația F1. Procedand la
autopolenizarea hibrizilor din generația F1 a obținut cea
de-a doua generație (F2), care a constat dintr-un total de 1064
de plante, dintre care 277 au fost pitice și 787 au fost inalte. Raportul
caracterelor planta inalta : planta pitica in generația F2
a fost de 2,84 : 1, prin rotunjire obținandu-se un raport de 3 : 1. Mendel
a obținut aceleași rezultate și atunci cand a fost inversata
inalțimea parinților, demonstrand ca nu are
importanța care planta este folosita ca genitor matern sau
ca genitor patern.
De asemenea, prin hibridarea intre o
planta homozigota cu semințe netede (caracter datorat
prezentei amidonului) cu o planta homozigota, cu semințe
zbarcite (caracter datorat prezentei dextrinei), Mendel a obtinut in
prima generatie numai plante cu semințe netede. Acest caracter a fost
denumit dominant, in timp ce caracterul semințe zbarcite, care nu
a aparut in F1, a fost denumit recesiv. Prin
autopolenizarea plantelor din prima generatie, a obtinut in
generatia a doua atat plante cu semințe netede, cat si plante cu
semințe zbarcite, proportia dintre caracterul dominant si cel
recesiv fiind de 3:1.
Raportul de 3 : 1 intre caracterul dominant
și cel recesiv a fost obținut și pentru celelalte perechi de
caractere studiate, dupa cum este prezentat in tabelul
X.
Deci, in fiecare experiment Mendel a observat
ca un caracter al unui genitor este dominant la hibrizii F1,
iar dupa autopolenizarea acestora cele doua caractere parentale
segrega intr-o proporție de 3 : 1.
|
Caracterul studiat
|
Caractere
|
Proporția
D: r
|
|
Dominant
(D)
|
Recesiv
(r)
|
Dominant
(D)
|
Recesiv
(r)
|
|
Talia plantei
|
Inalta
|
Pitica
|
|
|
|
|
Aspectul semințelor
|
Neteda
|
Zbarcita
|
|
|
:1,0104
|
|
Culoarea cojii boabelor
|
Colorata
|
Alba
|
|
|
|
|
Culoarea cotiledoanelor
|
Galbena
|
Verde
|
|
|
|
|
Consistenta
pastailor
|
Tare
|
Moale
|
|
|
|
|
Culoarea pastailor
|
Verde
|
Galbena
|
|
|
|
|
Dispozitia florilor
|
Axilara
|
Terminala
|
|
|
|
|
Total pentru toate insusirile
|
|
|
|
|
|
Prin autopolenizarea
plantelor din cea de-a doua generație, Mendel a observat ca la o
treime din plante (inalte, homozigote), in urma segregarii factorilor
ereditari s-au format exclusiv plante inalte, la doua treimi dintre plante
(inalte, heterozigote) factorii ereditari au segregat in dominanți și
recesivi, in raport de 3 : 1 și in descendența apar doar plante
inalte (homozigote), in timp ce prin autopolenizarea plantelor pitice
(homozigote), in descendența apar doar plante pitice (homozigote).
Segregarea genelor: Pe baza rezultatelor obținute
in experimentele de hibridare realizate la mazare, Mendel a concluzionat
ca fiecare genitor contribuie cu un factor ereditar pentru
un caracter la hibrizii din F1. Astfel, hibrizii din prima
generație au cate doi factori ereditari pentru fiecare caracter, care se
separa unul de celalalt atunci cand acești hibrizi formeaza
gameți. Segregarea genelor a devenit astfel PRIMA LEGE A LUI MENDEL
și a fost denumita LEGEA SEGREGARII FACTORILOR EREDITARI sau
LEGEA PURITAȚII GAMEȚILOR.
Gametii sunt intotdeauna puri
din punct de vedere genetic, adica nu contin decat unul dintre factorii
ereditari. Prin combinarea probabilistica a acestor gameti
rezulta in generatia a doua, in urma fenomenului de segregare, un
raport de 3 dominant : 1 recesiv.
Aceasta lege
poate fi reprezentata schematic astfel:
Gentori: Planta
inalta (homozigota) x Planta pitica (homozigota)
Factori ereditari: TT tt
Gameti: T t
Hibrizi F1: Planta
inalta (Tt) x Planta inalta (Tt)
Gameti: T,
t T, t
F2: 3
plante inalte : 1 planta pitica
TT, Tt, Tt : tt
MENDEL a denumit plantele, care contin
un singur tip de factor ereditar homozigote, iar cele care
contin ambele tipuri, heterozigote. La indivizii
heterozigoti nu se manifesta decat unul dintre factorii ereditari,
cel dominant (bob neted), celalalt ramanand in stare ascunsa (bob
zbarcit). In acest fel, G. MENDEL a reusit sa descopere deosebirea
dintre structura genetica a organismelor si
infatisarea lor, notiune care mai tarziu a fost
denumita de JOHANSEN genotip si respectiv fenotip. Genotipul
reprezinta totalitatea genelor continute de un organism. Fenotipul
reprezinta suma insusirilor morfologice, fiziologice, biochimice
si de comportament a un organism ca rezultanta a interactiunii
dintre genotip si mediu. Din exemplul anterior se remarca faptul
ca plantele cu acelasi fenotip (75% plante inalte), prezinta
genotipuri diferite: 25% sunt homozigote si 50% sunt heterozigote.
Dupa opt ani de investigații
detaliate, in anul 1866, Mendel a publicat rezultatele obținute intr-o
carte intitulata "Experiments in Plant Hybridisation", in proceedings of
the Brunn Natural History Society, insa lucrarea sa nu a primit nicio
atenție timp de 34 ani, cand, in anul 1900 De Vries, Correns și
Tschermak, lucrand independent au confirmat rezultatele lui Mendel.
Termeni utilizați:
Dominant, recesiv: Atunci cand in hibridarea intre
doua varietați homozigote, un caracter manifestat de unul dintre
cei doi genitori se exprima nemodificat la hibrizii din prima
generație este denumit cu termenul dominant. Caracterul care
nu apare in F1, dar apare in F2, este denumit cu termenul
recesiv.
Alele: Factorii ereditari care determina
expresii contrastante ale unui caracter se numesc alele sau alelomorfe. In
exemplul de mai sus, caracterul luat in studiu este inalțimea
plantei, iar factorii ereditari T și t, care controleaza dezvoltarea
unei plante inalte și, respectiv, a unei plante pitice, sunt alele.
Factorii ereditari au fost denumiți gene, in anul 1909, de
catre biologul danez Johannsen, astfel ca genele T și t sunt
gene alele sau alelomorfe.
Test cross (backcross sau
retroincrucișarea): Pentru verificarea rezultatelor obținute Mendel a realizat
hibridari intre plante de mazare pitice și inalte, in totalitate
homozigote, obținand in generația F1 numai plante de talie
inalta, unele cu genotip homozigot TT, iar altele cu genotip
heterozigot Tt. Astfel, la un hibrid Tt, factorii ereditari T și t
segrega la formarea gameților intr-un raport de 1 : 1, ceea ce
inseamna ca 50% dintre gameții unui hibrid heterozigot F1
poarta factorul T și 50% poarta factorul t. Mendel
a incrucișat un hibrid heterozigot (Tt) cu o varietate pitica
homozigota (tt), ai carei gameți erau purtatori ai
factorului ereditar recesiv t. Intr-o proproție de 50%, fiecare gamet al
genitorului homozigot (pentru factorul recesiv t) are șansa de a se combina
cu un gamet putator al factorului dominant T și intr-o proporție
de 50% șansa de a se combina cu un gamet purtator al factorului
recesiv t, formați de genitorul heterozigot. In consecința, 50%
dintre descendenți au fost inalți din punct de vedere fenotipic
și heterozigoți (Tt) din punct de vedere genotipic, in timp ce 50%
dintre descendenți au fost de talie pitica din punct de vedere
fenotipic și homozigoți (tt) din punct de vedere genotipic, dupa
cum reiese din urmatoarea schema:
Genitori: Tt
x tt
Gameți: T
(50%), t (50%) t (100%)
Test Cross: Planta
inalta : Planta pitica (1 : 1)
Descendența: Tt (50%) tt (50%)
Corectitudinea interpretarii
rezultatelor sale a fost confirmata de rezultatele acestor hibridari,
care erau similare cu așteptarile teoretice. Incrucișarea unui
individ (al carui genotip pentru un anumit caracter este necunoscut) cu un
altul dublu recesiv, pentru a determina și verifica genotipul primului
genitor, se numește testcross sau backcross.
Cu scopul de a determina genotipurile
hibrizilor din generația F2, Mendel a permis autopolenizarea
acestora, observand ca plantele de talie inalta, homozigote, au
format in generația F3, exclusiv descendenți de talie
inalta, al caror genotip este TT. In mod similar, prin
autopolenizarea plantelor pitice, homozigote s-au obținut exclusiv
descendenți de talie pitica, al caror genotip era tt. Prin
autopolenizarea heterozigoților F2, au fost obținuți
descendenți de talie inalta și de talie pitica, in raport
de 3 : 1, in mod similar cu hibrizii F1, ceea ce inseamna
ca genotipul lor era Tt.
Este important de subliniat faptul ca,
genotipul se noteaza utilizand doi factori ereditari (ex. TT, tt, Tt),
ceea ce este in acord cu ipoteza lui Mendel, conform careia fiecare
genitor are doi factori ereditari pentru fiecare caracter. De asemenea,
prezența a doi factori ereditari pentru fiecare caracter poate fi
explicata și din punct de vedere citologic. Astfel, la plante și
la animale cromosomii somatici exista sub forma perechilor de omologi, in
fiecare pereche un cromosom fiind de origine paterna, iar celalalt de
origine materna. Gena reprezinta o secvența din
macromolecula de ADN cromosomial, care deține informația
genetica necesara petru codificarea unui anumit caracter. Presupunand
ca o oarecare gena A, ocupa un anumit locus pe un cromosom, pe
cromosomul omolog, in locusul corespunzator se gasește gena
alela a, care codifica același caracter (ca și gena A). In
cursul diviziunii reducționale a meiozei (metafaza I), cromosomii omologi se
separa și migreaza spre poli opuși și ca urmare,
genele (factorii ereditari) segrega una de alta și ajung in
gameți diferiți.
Dihibridarea si legea
segregarii independente a perechilor de caractere
Dihibridarea consta in hibridarea a doi
indivizi, care se deosebesc prin doua perechi de caractere.
Mendel a
realizat hibridarea intre o planta de mazare cu bobul neted si
culoare galbena, reprezentand genitorul patern, și o planta de
mazare cu bobul zbarcit si de culoare verde, reprezentand genitorul
matern. In prima generatie hibrida, toate plantele aveau boabe netede
si de culoare galbena, indicand faptul ca, bobul neted și
culoarea galbena a cotiledoanelor sunt caractere dominante, comparativ cu
bobul zbarcit și culoarea verde a cotiledoanelor, care sunt caractere
recesive. Prin autopolenizarea plantelor din prima generatie s-a
obtinut generatia a doua (F2) in care descendenții
prezentau caracterele genitorilor in diferite combinații, dupa cum
urmeaza: aproximativ 9/16 din plante aveau boabe netede si galbene,
3/16 zbarcite si galbene, 3/16 netede si verzi si 1/16 zbarcite
si verzi. In urma unei hibridari reciproce, in care genitorul matern
avea semințe netede si cu cotiledoanele de culoare galbena, iar
genitorul patern avea semințe zbarcite și cu cotiledonele de culoare
verde a condus la același rezultat.
In mod ipotetic, incrucisarea
unei forme de salcam cu flori albe (AA) si cu lujeri spinosi (SS)
caractere dominante, cu o forma cu flori roze (aa) si lujeri
nespinosi (ss) - caractere recesice - in prima generatie s-ar
obtine numai exemplare cu flori albe si lujeri spinosi.
In F2, la arborii
obtinuti prin autofecundarea hibrizilor F1 s-ar ajunge la
urmatoarea repartitie a caracterelor: 9/16 plante cu flori albe
si lujeri spinosi, 3/16 plante cu flori albe si lujeri
nespinosi, 3/16 plante cu flori roze si lujeri spinosi si
1/16 plante cu flori roze si lujeri spinosi (STANESCU, 1982).
Mendel a aplicat principiul
monohibridarii și a considerat genitorii homozigoți, cu ambele
caractere dominante, respectiv RRYY genitorul cu semințe netede si cu
cotiledoane de culoare galbena și rryy genitorul cu semințe
zbarcite și cu cotiledonele de culoare verde. Avand in vedere ca
fiecare caracter este determinat de doi factori ereditari, in cazul
dihibridarii se iau in considerare patru factori ereditari pentru fiecare
genitor, ceea ce insemna ca hibrizii F1 trebuie sa
fie RrYy.
Raportul de segregare de 9 : 3 : 3 : 1
in generația F2, a fost explicat de catre Mendel printr-o
segregare independenta a perechilor de caractere, semințe netede
și semințe zbarcite pe de o parte, respectiv semințe cu
cotiledoane galbene și semințe cu cotiledoane verzi, pe de alta
parte. Astfel, cand hibrizii F1 vor forma gameți, exista
probabilitatea ca jumatate dintre aceștia sa aiba factorul
ereditar R și jumatate factorul ereditar r. O probabilitate
similara se intalnește și in cazul gameților conținand
factorii ereditari Y și y. Prin urmare, probabilitatea ca factorii
ereditari R și Y sa ajunga in același gamet este de 25%,
probabilitatea fiind aceeași și pentru factorii ereditari R și
y, r și Y, precum și, y și r. Deci, gameții conținand
factorii ereditari RY, Ry, rY și ry trebuie sa se formeze in
proporții egale.
Pentru determinarea descendenței
in cea de-a doua generație, este utila alcatuirea unui
patrat Punnett (similar unei table de șah), in care gameții
produși de unul dintre genitori sunt plasați in partea de sus a
tablei, iar gameții produși de celalalt genitor in partea
laterala. In cele 16 patrațele ale tablei se inscriu
posibilele combinații dintre gameții feminini și gameții
masculini, in procesul fecundarii (Fig. X).
Citirea fenotipurilor diferiților descendenți, indica un raport
de segregare de 9 : 3 : 3 : 1, așa cum a observat și Mendel.
Genitorii: RRYY x rryy
Gameții: RY
ry
F1: RrYy x RrYy
Gameții: RY, Ry, rY, ry RY, Ry, rY, ry
F2:
|
RY
|
Ry
|
rY
|
ry
|
|
RY
|
RRYY
|
RRYy
|
RrYY
|
RrYy
|
|
Ry
|
RRYy
|
RRyy
|
RrYy
|
Rryy
|
|
rY
|
RrYY
|
RrYy
|
rrYY
|
rrYy
|
|
ry
|
RrYy
|
Rryy
|
rrYy
|
rryy
|
Ca și in cazul
monohibridarii, Mendel a verificat rezultatele obținute realizand o
retroincrucișare (test cross sau backcross) intre hibrizii F1,
heterozigoți pentru ambele caractere (RrYy) cu genitorul dublu recesiv
(rryy), care formeaza un singur tip de gameți, și anume ry.
Hibridul RrYy produce patru tipuri de
gameți, respectiv RY, Ry, rY, ry, cu o frecvența egala,
ceea ce inseamna ca daca in procesul fecundarii acești
gameți se unesc cu gameții ry ai genitorului recesiv, descendența
trebuie sa manifeste toate cele patru combinații ale caracterelor in
proporții egale, așa cum a obținut Mendel, rezulate prezentate
in figura X.
F1: RrYy x rryy
Gameții: RY, Ry, rY, ry ry
|
RY
|
Ry
|
rY
|
ry
|
|
ry
|
RrYy
|
Rryy
|
rrYy
|
rryy
|
Mendel a largit aria de
observații realizand trihibridari in care au fost implicați
genitori care se deosebeau prin trei perechi de caractere: aspectul
semințelor: netede (S) - zbarcite (s), culoarea cotiledoanelor:
galbena (Y) - verde (y) și culoarea florilor: violet (V) - alba
(v). In generația F1 a obținut hibrizi care prezentau
toate caracterele dominante (SsYyVv), fiind heterozigoți pentru toate
caracterele luate in studiu. Hibrizii F1 formeaza opt tipuri de
gameti masculini si opt tipuri de gameti feminini, prin a
caror unire pe baza de probabilitate se formeaza in
generația F2, 64 de combinatii si 8 tipuri de
organisme:
27/64 cu trei caractere dominante
(SYV);
9/64 cu doua caractere
dominante si unul recesiv (SYv);
9/64 cu doua caractere
dominante si unul recesiv (SyV);
9/64 cu doua caractere dominante
si unul recesiv (sYV);
3/64 cu un caracter dominant
si doua recesive (Syv);
3/64 cu un caracter dominant
si doua recesive (sYv);
3/64 cu un caracter dominant
si doua recesive (syV);
1/64 cu trei caractere recesive
(syv).
In cazul trihibridarii, ca și in
cazul tuturor hibridarilor multiple, pentru determinarea descendenței
in generația F2 este utila metoda multiplicarii,
comparativ cu alcatuirea unui patrat Punnett.
Metoda multiplicarii presupune
calcularea produsului dintre sumele perechilor de factori ereditari, in
raportul obținut conform monohibridarii, respectiv 3 :1., adica:
(3S + 1s) x (3Y + 1y) x (3V + 1v). Aceasta multiplicare poate fi
realizata intr-o maniera pas cu pas: intr-o prima etapa se
realizeaza multiplicarea (3S + 1s) x (3Y + 1y) = 9SY + 3Sy + 3sY + 1sy,
iar intr-o a doua etapa acest rezultat se inmulțește cu raportul
se segregare al celei de-a perechi de factori ereditari, respectiv (3V + 1v).
Astfel, (9SY + 3Sy + 3sY + 1sy) x (3V + 1v) = 27SYV + 9SYv + 9SyV + 3Syv + 9sYV
+ 3sYv + 3syV + 1syv. Deci, raportul de segregare este 27 : 9 : 9 : 9 : 3 : 3 :
3 :1, conform rezultatelor obținute de Mendel.
In acest fel, Mendel a ajuns la concluzia
ca, la incrucisarea intre organisme ce se deosebesc prin doua
sau mai multe perechi de caractere are loc fenomenul segregarii independente a
perechilor de caractere. El a considerat, de asemenea, ca
ereditatea unei perechi de caractere este independenta de ereditatea
celeilalte perechi de caractere, astfel ca si segregarea are loc
independent. Daca in exemplul cu dihibridarea, se studiaza segregarea
perechii de caractere semințe netede si semințe zbarcite,
fara sa se tina seama de culoarea bobului
(galbena sau verde), atunci se obtine acelasi raport de
segregare de 3:1 intre cele doua caractere. Acelasi lucru se
intampla si in cazul culorii cotiledoanelor semințelor. Aceasta
este segregarea de tip Pisum
denumita astfel deoarece a fost descoperita la mazare. In
conformitate cu acest tip de segregare, in F2 heterozigotii de
tip Aa sunt identici fenotipic cu homozigotii AA.
Interactiunea genelor
Raportul fenotipic obținut de Mendel in
urma experimentelor de hibridare realizate la mazare, a demonstrat ca
o gena controleaza un anumit caracter, iar dintre cele doua
alele ale unei gene, una este complet dominanta asupra
celeilalte. Ca o consecința a acestui fapt, heterozigoții au un
fenotip identic cu homozigoții, cu ambele gene dominante. La scurt timp
dupa redescoperirea legilor mendeliene, o serie de experimente au
evidențiat faptul ca uneori o gena nu are un efect individual,
ci mai multe gene interacționeaza pentru a produce un nou fenotip.
Intr-unul dintre primele cazuri raportate de
Kölreuter, heterozigoții prezentau un fenotip intermediar intre
fenotipurile genitorilor, fenomen care poarta denumirea de dominanța
incompleta sau ereditate intermediara. In cazul
codominanței, fenotipurile genitorilor se manifesta in
proporții egale la heterozigoți, iar in cazul epistaziei o
gena inhiba activitatea unei alte gene, dintr-un anumit locus. In
alte situații, o gena modifica efectul unei alte gene, in sensul
diminuarii sau amplificarii expresiei celei de-a doua gene. Astfel de
gene poarta denumirea de gene modificatoare.
Interacțiunea dintre gene, reprezinta activitatea comuna a mai
multor gene, care are un efect ce nu poate fi determinat de nicio gena, in
mod independent. Astfel de gene se numesc complementare. De
asemenea, exista gene care copiaza activitatea altor gene, altfel
spus, determina un efect similar, acestea fiind denumite izogene
(gene duplicate). Nu in ultimul rand, exista gene care cauzeaza
moartea organismului, acestea fiind cunoscute sub denumirea de gene
letale.
Deci, abaterile de la legile mendeliene ale
ereditatii apar in urma mecanismelor de interactiune a genelor,
ce genereaza fenomene de dominanta completa,
incompleta, letalitate s.a. Raportul de segregare genotipic in F2
este acelasi, dar in functie de relatiile existente intre genele
alele sau nealele, aceste constitutii genotipice determina fenotipuri
diferite.
Dominanta incompleta (semidominanta)
Daca, in cazul dominantei
complete, organismele homozigote (AA) prezinta acelasi fenotip cu
cele heterozigote (Aa), in cazul dominanței incomplete fenotipul
organismelor ce contin genele alele in stare heterozigota se deosebeste
de cel al organismelor ce contin genele respective in stare
homozigota (deci AA≠Aa). Fenomenul mentionat este cunoscut sub
denumirea de semidominanta sau dominanta
incompleta.
Experimentele de monohibridare realizate la Mirabilis jalapa, intre doua varietați
distincte prin culoare florilor, au condus la descoperirea fenomenului de
semidominanța, de catre Correns, in anul 1912. Incrucisand o planta cu flori rosii (RR), cu una cu
flori albe (rr), a obtinut in generatia F1 plante cu flori
de o culoare intermediara, roz (Rr). Indivizii heterozigoti (Rr)
manifesta caractere intermediare intre fenotipurile genitorilor,
datorita dominantei incomplete sau semidominantei genei R - dominanta asupra genei r - recesiva. Prin autopolenizarea
plantelor hibride F1, in F2 s-au obtinut atat plante
cu flori rosii si albe, dar si plante cu flori roz. Raportul
fenotipic este identic cu cel genotipic 1RR (flori rosii) : 2Rr (flori
roz) : 1rr (flori albe). Același raport de segregare fenotipic se
obține și in cazul monohibridarii dintre varietatea cu flori
roșii și varietate cu flori albe, la specia Antirrhinum majus.
Dominanța incompleta este
explicata prin absența dominanței atat in ceea ce privește
caracterul flori albe, cat și in cazul caracterului flori roșii, dar
fiecare alela are o anumita influența in formarea culorii,
astfel incat hibrizii au flori de culoare roz.
Acest tip de ereditate
intermediara a fost observata la multe alte specii de pante, dar
și la animale.
La Zea
mays, din incrucisarea unei varietati cu boabe albastre cu
una cu boabe galbene au rezultat in F1 plante cu boabe violete,
culoare intermediara intre genitori. In F2 s-a produs
segregarea astfel:1 albastru : 2 violet : 1 galben. Acest tip de segregare (cu
un raport fenotipic de 1 : 2 : 1), in care organismele heterozigote prezinta
fenomenul dominantei incomplete, este cunoscut și sub denumirea de segregare
de tip Zea, pentru a-l deosebi de segregarea de tip Pisum (cu un
raport fenotipic de 3 : 1) observata de Mendel.
Dominanta incompleta
intervine si in cazul raporturilor de segregare la arbori, fenomenul
actionand probabil, in cazul unor caractere cum sunt forma acuminata
a solzilor conurilor de molid, in raport cu forma rotunjita sau culoarea
roscata a ritidomului, la pinul silvestru, in raport cu culoarea
cenusie-inchisa a ritidomului s.a.
La pasari fenomenul de
dominanta incompleta a fost observat la gainile de
Andaluzia, cu penaj de culoare albastruie. Ele provin din
incrucisarea unei rase de gaini cu penajul negru cu una cu penajul
alb. In F1 toate pasarile au penaj albastru. In F2
se produce segregarea astfel: 25% penaj negru, 25% penaj alb si 50% penaj
albastru. Aceasta inseamna ca gainile de Andaluzia cu penaj
albastru sunt heterozigote, fapt pentru care ele segrega mereu,
neputandu-se crea o rasa de gaini de Andaluzia.
Codominanta
Codominanta determina
aparitia la indivizii heterozigoti pentru genele dominante a unui
fenotip nou, comparativ cu indivizii homozigoti.
La indivizii din populația
umana, sistemul OAB al grupelor de sange este controlat de alele multiple
ale genei I, fiecare alela codificand un antigen diferit. De altfel, la
indivizii umani au fost identificate patru grupe de sange, desemnate: A, B, AB
și O. Indivizii cu grupa de sange O nu au nici un antigen, in timp ce la
heterozigoții cu grupa de sange AB prezinta ambele antigene,
respectiv A și B.
Genele IA si IB sunt dominante asupra genei i, iar impreuna sunt codominante, adica determina un fenotip nou - grupa
de sange AB. Ca urmare, indivizii pot fi fenotipic si genotipic de mai
multe tipuri (Tabel X).
Indivizii heterozigoți cu grupa
de sange AB prezinta in genotipul lor ambele alele dominante IA
și IB, care se exprima cu intensitate egala pentru a
determina formarea distincta a antigenelor A și B.
Genotipul heterozigot LALB se
manifesta prin grupa de sange AB. In mod similar, in sistemul MN al
grupelor de sange, alele M și N sunt codominante determinand apariția
grupei de sange MN la heterozigoți.
|
Fenotip
|
Genotip
|
|
ll
|
|
A
|
LALA
LAl
|
|
B
|
LBLB
LBl
|
|
AB
|
LALB
|
Tabel X.
Schema grupelor de sange din sistemul AB0
Pentru realizarea unor transfuzii
sangvine trebuie cunoscute grupele de sange ale donatorului si
receptorului. Cunoasterea modului cum se mostenesc grupele sangvine
are importanta si in stabilirea paternitatii.
De asemenea, interactiunea dintre
doua sau mai multe gene nealele,de asemenea dominante, prezente
impreuna, determina aparitia unui nou fenotip.
In anul 1900, Bateson și
colaboratorii au studiat modul de transmitere ereditara a formei crestei
la gaini, luand in considerare cele patru tipuri de creste intalnite,
respectiv: trandafir, bob de mazare, nuca și simpla.
Intr-un prim experiment de hibridare realizat intre gaini cu creasta
in forma de trandafir și gaini cu creasta simpla,
toți hibrizii F1 au prezentat creasta in forma de
trandafir, iar prin consangvinizare s-a obținut in cea de-a doua
generație un raport fenotipic de 3 : 1. In cel de-al doilea experiment de
hibridare intre gaini cu creasta bob de mazare și
gaini cu creasta simpla, hibrizii F1 au prezentat in
exclusivitate creasta bob de mazare, iar in F2 s-a
obținut segregarea caracterelor creasta bob de mazare și
creasta simpla in raport de 3 : 1. Deci, gena pentru creasta bob
de mazare este dominanta asupra genei care codifica caracterul
creasta simpla.
In mod surprinzator, in urma incrucișarii
dintre o gaina cu creasta trandafir și o gaina cu
creasta bob de mazare, in generația F1 toți
descendenții au avut creasta tip nuca. Prin consangvinizarea
descendenților cu creasta tip nuca din generația F1,
s-a obținut in generația F2 pui cu toate tipurile de
creste: trandafir, bob de mazare, nuca și simpla, in raport
de 9 : 3 : 3 : 1. Avand in vedere ca acest raport de segregare este
caracteristic dihibridarii, se poate concluziona ca creste de tip
trandafir și de tip bob de mazare sunt codificate de doua
perechi de gene.
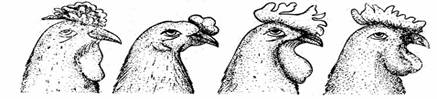
Trandafir Mazarata Nuca Simpla
Apariția puilor cu creasta
tip nuca in generația F1, in urma incrucișarii
dintre un genitor cu creasta tip trandafir și unul cu creasta
tip bob de mazare, indica faptul ca, independent genele
dominante P și R sunt responsabile de apariția caracterului
creasta tip nuca.
Atunci cand sunt prezente
impreuna in zigot, genele P și R interacționeaza in
formarea crestei tip nuca. Cand sunt prezente separat, determina fie
creasta de tip trandafir, fie creasta de tip bob de mazare.
Alele recesive pentru creste de tip trandafir și de tip bob de
mazare, determina cel de-al patrulea tip de creasta, respectiv
tipul simplu.
Genitori:
Gaina cu creasta trandafir
x Gaina cu
creasta mazarata
RRpp rrPP
Gameți: Rp rP
F1:
Pui cu creasta
nuca x Pui
creasta nuca
RrPp x RrPp
Gameți: RP, Rp, rP, rp RP, Rp, rP, rp
|
RP
|
Rp
|
rP
|
rp
|
|
RP
|
RRPP
(creasta tip nuca)
|
RRPp
(creasta tip nuca)
|
RrPP
(creasta tip nuca)
|
RrPp
(creasta tip nuca)
|
|
Rp
|
RRPp
(creasta tip nuca)
|
RRpp
(creasta tip trandafir)
|
RrPp
(creasta tip nuca)
|
Rrpp
(creasta tip trandafir)
|
|
rP
|
RrPP
(creasta tip nuca)
|
RrPp
(creasta tip nuca)
|
rrPP
(creasta tip bob de mazare)
|
rrPp
(creasta tip bob de mazare)
|
|
rp
|
RrPp
(creasta tip nuca)
|
Rrpp
(creasta tip trandafir)
|
rrPp
(creasta tip bob de mazare)
|
rrpp
(creasta simpla)
|
Epistazia
Epistazia consta in mascarea
expresiei fenotipice a unei gene numita hipostatica, de catre o
alta gena nealela numita epistatica.
Epistazia poate fi de dominanta
sau de recesivitate, dupa cum gena epistatica este dominanta sau
recesiva.
A. Epistazia de dominanta a fost evidentiata de Bateson
si Punnet, in experientele lor privind transmiterea colorata a
penajului la gaini. Culoarea penajului este determinata de doua
gene nealele.
Experimentele de hibridare realizate intre
rasele de gaini Leghorn și Wyandotte, au evidențiat faptul
ca, gena care determina culoarea alba a penajului la rasa
Leghorn este dominanta asupra genei care determina penajul colorat,
iar gena care determina culoarea alba a penajului la rasa Wyandotte
este recesiva fața de gena care determina penajul colorat.
Deci gena care determina culoarea alba a penajului la rasa Leghorn
este diferita de gena care determina culoarea alba a penajului
la rasa Wyandotte.
Prin incrucișarea dintre o
gaina din rasa Leghorn cu penajul alb și una din rasa Wyandotte
cu penajul alb, s-a obținut in F1 numai gaini cu penajul
alb, cu mici pete de culoare mai inchisa. Prin autofecundarea acestora, in
F2 s-a produs segregarea genelor intr-un raport de 13 gaini cu
penajul alb : 3 gaini cu penajul colorat. Rezultatul este explicat prin
existența a doua gene dominante la gainile cu penajul alb din
rasa Leghorn, dupa cum este prezentat in urmatoarea schema:
Genitorii:
Rasa alba Leghorn x Rasa alba Wyandotte
IICC iicc
Gameții:
IC
ic
F1: Gaini cu penajul alb, cu
mici pete de culoare inchisa
IiCc x IiCc
F2:
|
IC
|
Ic
|
iC
|
ic
|
|
IC
|
IICC
|
IICc
|
IiCC
|
IiCc
|
|
Ic
|
IICc
|
IIcc
|
IiCC
|
Iicc
|
|
iC
|
IiCC
|
IiCc
|
iiCC
|
iiCc
|
|
ic
|
IiCc
|
Iicc
|
iiCc
|
iicc
|
Analiza celor 16 fenotipuri si
genotipuri la hirbizii F2 a indicat faptul ca doar 3 dintre
cele 16 genotipuri, respectiv iiCC, iiCc, iiCc au determinat penajul colorat al
gainilor.
Evident, gainile Leghorn cu penajul
alb, prezinta gena I, care in stare homozigota (II) sau
heterozigota (Ii) inhiba sau suprima gena C pentru penajul
colorat. Gena C se exprima atunci cand gene pentru culoarea alba se
gasește in stare dublu recesiva (ii). Gena I se numește
epistatica, iar gena C se numește hipostatica.
Acesta este un exeplu de epistazie de
dominanța, deoarece chiar și numai o gena dominanta
(I) este capabila sa inhibe sau sa suprime gena C pentru penajul
colorat.
B. Epistazia de recesivitate. Actiunea unei gene recesive in stare
homozigota, gena epistatica, inhiba manifestarea altei gene
nealele, in stare homozigota sau heterozigota, gena hipostatica.
Culoarea blanii la caini este
controlata de doua gene: cc (gena epistatica), determina
culoarea alba; B - (gena hipostatica), care in absenta cc,
determina culoarea neagra; bb (gena hipostatica), in
absenta cc, determina culoarea maron. In urma incrucisarii
intre doua rase de caini, una homozigota cu blana alba, BBcc,
si alta homozigota cu blana maron, bbCC, se obtin in F1
numai indivizi cu blana neagra, BbCc. In F2 se obtine
raportul de segregare fenotipica: 9 blana neagra: 4 blana
alba: 3 blana maro. .
|
Genitorii:
|
BBcc
|
|
bbCC
|
|
(alb)
|
|
(maron)
|
|
F1:
|
|
BbCc
|
|
|
|
|
(negru)
|
|
|
F2
|
|
BC
|
Bc
|
bC
|
bc
|
|
BC
|
BBCC
negru
|
BBCc
negru
|
BbCC
negru
|
BcCc
negru
|
|
Bc
|
BBCc
negru
|
BBcc
alb
|
BbCc
negru
|
Bbcc
alb
|
|
bC
|
BbCC
negru
|
BbCc
negru
|
bbCC
maron
|
bbCc
maron
|
|
bc
|
BbCc
negru
|
Bbcc
alb
|
bbCc
maron
|
bbcc
alb
|
| | | | | | | | |
Figura X. Fenomenul de epistazie de recesivitate,
in transmiterea culorii blanii la caini (raport de segregare 9 blana
neagra: 4 blana alba: 3 blana maro).
Epistazia si grupele de sange la
om. In sistemul ABO al grupelor de sange la om,
indivizii cu grupa de sange A, prezinta pe suprafața hematiilor
antigenul A, indivizii cu grupa de sange B prezinta pe suprafața
hematiilor antigenul B, indivizii cu grupa de sange AB, prezinta pe
suprafața hematiilor ambele antigene A și B, iar indivizii cu grupa
de sange O nu au niciun antigen. Antigenele sunt controlate de o gena
autosomala I (izohemaglutinina), care prezinta mai multe alele.
Astfel, IA controleaza antigena A, IB
controleaza antigena B, iar alela recesiva i nu determina
producerea unei antigene, rezulștand astfel fenotipul O.
Aglutininele numite și anticorpi,
prezente in serul sangvin al indivizilor umani se gasesc intr-o
relație precisa cu antigenele de pe hematii. Serul sangvin al unui
individ cu grupa de sange A are anticorpi impotriva antigenelor B, in timp ce
serul sangvin al unui individ cu grupa de sange B are anticorpi impotriva
antigenelor A. La persoanele cu grupa de sange AB, care prezinta ambele
antigene A și B, serul sangvin nu conține anticorpi anti-A și
anti-B, iar persoanele cu grupa de sange O au ambele tipuri de anticorpi.
S-a descoperit ca pentru formarea
antigenelor A și B este responsabila o substanța
precursoare H, controlata de gena H. Astfel, la indivizii cu grupa de
sange A, sub controlul genei IA, precursorul H este convertit in
antigena A. Similar, la indivizii cu grupa de sange B, sub controlul genei IB,
precursorul H este convertit in antigena B. La indivizii cu grupa de sange O,
sub controlul genei recesive i, precursorul H nu este convertit intr-o
antigena, ramanand ca atare in sange. Precursorul H poate fi
aglutinat de catre anticorpii anti-H sau in prezența unor substanțe,
ca de exemplu extrctul din semințe de Ulex europeus. La indivizii
heterozigoți, cu grupa de sange A (IAi) sau B (IBi),
este aglutinat doar aprocentul de precursor H care nu este convertit intr-o
antigena. La indivizii cu grupa de sange A, precursorul AB nu este
aglutinat.
S-a constatat ca majoritatea oamenilor sunt homozigoți pentru gena H
(HH), puțini sunt heterozigoți (Hh) și foarte rar, gena este
prezenta in stare homozigota recesiva (hh), indivizii respectivi
fiind incapabili sa produca susbstanța precursoare H și in
consecința putand avea orice grupa de sange din sistemul ABO,
hematiile nefiind aglutinate nici de anti-A, nici de anti-B, nici de anti-H.
Descoperit pentru prima data in Bombay, India, la un individ homozigot
recesiv (hh), a fost denumit fenotipul Bombay, care a fost explicat prin
acțiunea epistatica a genei hh asupra genei I.
Complementaritatea genelor
Complementaritatea genelor
reprezinta, interactiunea dintre doua sau mai multe gene
nealele, care prezente impreuna, determina aparitia unui alt
fenotip, decat fenotipul determinat de fiecare gena in parte.
A. Complementaritatea de
dominanta a fost evidentiata
de BATESON si PUNNET (1908) la specia Lathyrus
odoratus. S-a remarcat ca in urma incrucisarii a doua
varietati de L. odoratus,
cu flori albe, in generatia F1 s-au obtinut plante cu
flori rosii, iar in generatia F2, raportul de segregare a
fost 9/16 plante cu flori rosii, 7/16 plante cu flori albe. Pentru
formarea antocianului este necesara prezenta concomitenta a
doua gene dominante (C si P). Fiecare genitor prezinta in stare
homozigota o gena dominanta si una recesiva (CCpp,
respectiv ccPP). In stare dominanta ambele gene sunt intalnite numai in F1
(CcPp) si 9/16 din plantele obtinute in F2 (C-P-).
Raportul fenotipic de segregare este de 9:7 (fig. 2.8.)
B. Complementaritatea de
recesivitate, a fost evidentiata de
SHULL in 1914, la specia Capsella bursa
pastoris (traista ciobanului). S-a remarcat ca forma capsulei plantei
este determinata de interactiunea dintre doua gene nealele. In
starea de recesivitate a ambelor gene, apare tipul cu capsula
ovoidala.
Daca una sau ambele gene se
afla in stare dominanta, apare tipul cu capsula
triunghiulara. Raportul genotipic de segregare in F2 este de
9/16 A-B (capsula triunghiulara):3/16 aaB (capsula
triunghiulara):3/16 A-bb (capsula triunghiulara):1/16 aabb
(capsula ovoidala), iar cel fenotipic de 15:1 (fig. 2.9).
Actiunea complementara a
genelor recesive se explica prin prezenta unor gene duplicate (izogene),
adica a unor gene similare ca actiune, care insa se transmit
independent. Planta Capsella bursa
pastoris fiind un tetraploid, are doua perechi din fiecare cromozom
omolog si ca atare poseda gene duplicate, care determina
actiunea de complementaritate recesiva.
Supradominanta
In acest caz, indivizii
heterozigoti depasesc in productivitate, fertilitate,
viabilitate etc., genitorii homozigoti, dominanti sau recesivi, AA<Aa>aa.
Fenomenul se numeste supradominanta
si el poate fi observat atat la plante cat si la animale. In procesul
de aparitie a supradominantei sunt de obicei, implicate mai multe
gene cu caracter aditiv. Fenomenul mai poarta denumirea de heterozis
monogenic.
La Drosophila
melanogaster, heterozigotii ww+, prezinta o cantitate
mai mare de pigmenti in ochi (sepiaterina), fata de genitorii
homozigoti w+w+ (ochi rosu-caramiziu)
si ww (ochi albi).
Pleiotropia
Prin fenomenul de pleiotropie se
intelege efectul fenotipic multiplu al unei singure gene.
Genele care au capacitatea de a
determina la acelasi organism doua sau mai multe caractere se numesc pleiotropice.
Fenomenul a fost observat de MENDEL,
care a demonstrat ca mazarea cu flori albe are boabe cu tegumentul
alb, cea cu flori colorate are tegumentul cenusiu sau brun.
NILLSON-EHLE a aratat ca o
anumita gena la ovaz modifica simultan forma aristelor
spicului, pilozitatea si fragilitatea tulpinilor.
Un exemplu caracteristic de
pleiotropie il ofera hibridul Nicotiana
silvestris Nicotiana tabacum, la care apar sase
caractere distincte, deschiderea carpelelor, transformarea ovulelor in carpele,
alungirea axei florale deasupra carpelelor, placentatia, cantitatea de
nectar si incretirea bazala a corolei, care sunt controlate de
aceeasi gena
Genele letale
Genele letale schimba raportul
mendelian de segregare, deoarece prezenta lor in stare homozigota
(dominanta sau recesiva) determina moartea indivizilor
purtatori, inainte de maturitatea sexuala.
Dupa gradul de manifestare,
efectul genelor letale poate fi:
gene letale, ce cauzeaza o
mortalitate de 100% a indivizilor purtatori;
gene semiletale, in acest caz 50%
dintre indivizii purtatori mor;
gene subvitale, care cauzeaza
moartea unui numar mai mic decat 50% dintre indivizii purtatori.
Notiunea de gena letala
a fost introdusa in stiinta de catre CUÉNOT (1911), in
urma cercetarilor intreprinse pe soareci de culoare galbena.
Acestia s-au dovedit a fi intotdeauna heterozigoti, deoarece prin
incrucisarea a doi soareci galbeni, nu se obtin niciodata
numai soareci galbeni, ci un amestec cu indivizi de alta culoare.
Raportul intre soarecii galbeni si cei de alta culoare este in
general de 2:1.
Daca se noteaza genele
pentru culoarea normala a blanii cu a si mutanta sa care determina culoarea galbena a
blanii cu Ay (yellow = galben), prin incrucisarea intre
organisme heterozigote ar trebui sa se obtina raportul de 3:1
intre indivizii galbeni si de alta culoare.
Aya Aya=AyAy+2Aya+aa
Daca se tine seama de
tipul AyAy, adica galben homozigot lipseste,
s-a dedus ca gena Ay este o alela mutanta
letala, care in stare homozigota duce la moartea organismului. S-a
constatat ca prin sacrificarea unei femele gestante, o parte a
soarecilor de culoare galbena mor in stadiu embrionar.
La plante au fost identificate
numeroase gene letale care in stare homozigota provoaca moartea
embrionilor, sau plantelor inainte de maturitate. La porumb se cunosc 15 gene
diferite care in stare homozigota blocheaza fotosinteza si
formarea clorofilei (plante albinotice), incapabile sa
supravietuiasca. Acelasi fenomen a fost semnalat si la Antirrhium majus.
Poligenia (polimeria)
Mai multe gene nealele, care segrega
independent, pot afecta acelasi caracter, expresia fenotipica
depinzand de numarul de gene dominante prezente.
NILSSON-EHLE a constat ca la grau, in
determinarea culorii bobului sunt implicate 2 (3) perechi de gene nealele,
notate cu R1, R2, (R3).
Culoarea bobului depinde de numarul de
gene dominante prezente si numarul de gene nealele (2 sau 3),
implicate. Din figura 2.14, rezulta ca intensitatea culorii boabelor
de grau, la indivizi rezultati in F2, este
proportionala cu numarul de gene in stare dominanta
prezente in genotipul lor. Indivizii cu 4 gene R (R1R1R2R2)
in genotipul lor, vor avea bobul rosu inchis, cei cu trei gene R, vor avea
bobul rosu, cei cu doua gene R, vor avea bobul rosietic, cei cu
o gena R, vor avea bobul rosu pal, iar cei cu ambele gene in stare
recesiva (r1r1r2r2), vor avea
bobul alb. Raportul de segregare in F1 este de 1:4:6:4:1,
formandu-se 5 grupe fenotipice.
|
Genitorii:
|
R1R1R2R2
|
|
r1r1r2r2
|
|
bob rosu inchis
|
|
bob alb
|
|
F1:
|
|
R1r1R2r2
|
|
|
|
|
bob rosietic
|
|
|
F2:
|
|
R1R2
|
R1r2
|
r1R2
|
r1r2
|
|
R1R2
|
R1R1R2R2
rosu inchis
|
R1R1R2r2
rosu
|
R1r1R2R2
rosu
|
R1r1R2r2
rosietic
|
|
R1r2
|
R1R1R2r2
rosu
|
R1R1r2r2
rosietic
|
R1r1R2r2
rosietic
|
R1r1r2r2
rosu pal
|
|
r1R2
|
R1r1R2R2
rosu
|
R1r1R2r2
rosietic
|
r1r1R2R2
rosietic
|
r1r1R2r2
rosu pal
|
|
r1r2
|
R1r1R2r2
rosietic
|
R1r1r2r2
rosu-pal
|
r1r1R2r2
rosu pal
|
r1r1r2r2
alb
|
| | | | | | | | |
Figura X. Fenomenul de poligenie (polimerie) in
transmiterea culorii cariopsei la grau (raport de segregare fenotipic 1
rosu inchis:4 rosu:6 rosietic:1 alb)
Transgresiunea
Mai multe gene nealele, determina in
generatiile de segregare F2-F5, la o parte din
descendenti, depasirea expresiei maximale si minimale a
unui caracter, fata de gradul de manifestare a acestui caracter la
genitori.
Prin incrucisarea unor iepuri, cu
blana de culoare inchisa (M1M1M2M2M3M3m4m4),
cu iepuri cu blana de culoare deschisa (m1m1m2m2m3m3M4M4),
in prima generatie s-au obtinut indivizi heterozigoti cu
blana de culoare intermediara (M1m1M2m2M3m3M4m4).
Din descendenta acestora, pe langa indivizi cu diferite nuante
ale blanii, apar si iepuri cu blana alba (m1m1m2m2m3m3m4m4),
transgresiune
negativa (inglobeaza toate genele recesive), precum si
iepuri cu blana de culoare neagra (M1M1M2M2M3M3M4M4),
transgresiune
pozitiva (inglobeaza toate genele dominante). Formele
transgresive pozitive pot fi utilizate ca genitori valorosi, in procesul
de ameliorare.
Abaterile reale de la raporturile mendeliene
Exceptiile
de la tipurile de segregare mendeliana prezentate anterior, sunt in fond
aparente, deoarece mecanismul separarii factorilor ereditari si al
formarii combinatiilor ramane de tip mendelian.
Exista insa si abateri
reale de la proportiile respective, avand o alta sursa,
cum ar fi segregarea preferentiala a cromozomilor, nesepararea
cromozomilor omologi in meioza, formarea nerandomizata
a zigotilor la fecundare.
Astfel, repartitia intamplatoare in
meioza a cromozomilor omologi la celulele fiice si formarea
gametilor cu cromozomi omologi si gene alele in raport de 1:1 sunt uneori
deranjate, producandu-se segregarea preferentiala a unor
cromozomi. In consecinta, in macrosporogeneza, la
plante, se intampla ca un cromozom sa se localizeze de
preferinta in oosfera, iar omologul lui in unul din
ceilalti nuclei ai sacului embrionar, neparticipand astfel la formarea
zigotului. Aberatii similare pot sa apara si la formarea
spermatiilor si a polenului si ele afecteaza proportiile in
care apar grupele de fenotipuri in descendenta.
Nondisjunctia cromozomilor in
meioza are
ca efect la prima diviziune aparitia unei celule haploide cu un cromozom
suplimentar si a celeilalte celule haploide cu un cromozom in minus.
Celulele sexuale care iau nastere vor produce astfel in procesul
fecundarii zigoti cu 2n+1 si 2n-1 cromozomi, deci cu alte caractere
si cu alta repartitie la descendenti decat in cazul normal.
Formarea nerandomizata a
zigotilor,
adica unirea gametilor in procesul fecundarii nu se
realizeaza in mod intamplator, ci dupa anumite preferinte
conditionate genetic, cum ar fi viabilitatea inegala a celulelor
sexuale mascule sau capacitatea de formare si maturizarea inegala a
stigmatului. Fecundarea nerandomizata duce, de asemenea, la modificarea
raporturilor de segregare mendeliene.

