Biologie
DINAMICA FONDULUI DE GENE INTR-O
POPULATIE EXPERIMENTALA DE DROSOPHILA
MELANOGASTER, SUB PRESIUNE SELECTIVA
CUPRINS:
Introducere
I. Istoric
I.1. Din istoricul cercetarilor
I.1.1. Din istoricul cercetarilor pe plan mondial
I.1.2. Din istoricul cercetarilor efectuate in Romania
II. Drosophila melanogaster -
caracterizare general-biologica
si ecologica
II.1. Incadrarea sistematica a speciei Drosophila melanogaster
II.2. Date morfologice
II.3. Ciclul de viata
II.4. Raspandire si ecologie
II.4.1. Habitatul natural si conditiile de laborator
II.4.2. Temperatura - factor fizic ce influenteaza dezvoltarea
speciei Drosophila melanogaster
II.4.3. Umiditatea - factor fizic ce influenteaza dezvoltarea
speciei Drosophila melanogaster
II.4.4. Prolificitatea - factor ce influenteaza dezvoltarea
speciei Drosophila melanogaster
II.4.5. Longevitatea - factor ce influenteaza dezvoltarea
speciei Drosophila melanogaster
II.5. Unii paraziti ai culturilor de Drosophila melanogaster
II.6. Determinismul sexelor la Drosophila
melanogaster
III. Elemente de genetica a populatiilor si evolutie
III.1. Definitia si caracterizarea populatiei
III.2. Factorii care determina compozitia si dinamica
genetica a populatiilor
IV. Scopul investigatiilor
V. Materiale si metode de lucru
VI. Rezultate si discutii
Concluzii
Bibliografie
INTRODUCERE
Genetica, termen provenit din grecescul gennao - "a naste", introdus in anul 1906 de catre W.
Bateson, este disciplina biologica ce se preocupa cu studiul
ereditatii, variabilitatii si determinismului
caracterelor.
Dintre toate
ramurile biologiei, ea a cunoscut pana in zilele noastre cea mai
accelerata ascensiune, numerosi savanti consacrandu-se unei
munci perseverente si pasionante, efectuand studii compexe in domeniul
ereditatii si variabilitatii organismelor,
raspunzand unor intrebari care, de milenii, au preocupat omenirea. De
la procesele ce guverneaza dezvoltarea plantelor, animalelor si microorganismelor,
pana la ameliorarea soiurilor de plante si animale, de la structura
genei, pana la terapia genetica a cancerului, genetica a fost chemata
sa gaseasca raspunsuri, solutii, explicatii pe care
nici o alta disciplina bilogica nu le-a putut oferi.
A fost nevoie
de conlucrarea stransa cu numeroase alte ramuri ale biologiei, ca:
fiziologia, biochimia, biofizica, citologia, ecologia, microbiologia,
virusologia, etc. Aceasta conlucrare a permis geneticii o dezvoltare
furtunoasa, din ea separandu-se alte ramuri noi care studiaza
ereditatea la diferite nivele, cu ajutorul unor metode specifice. Astfel,
descoperirea procesului de diviziune celulara si aprofundarea
studiului componentelor celulare, mai ales al compexului nucleu - citoplasma,
a realizat o conexiune puternica intre citologie si genetica,
conexiune ce a dat mai tarziu nastere citogeneticii.
Descoperirea
energiei atomice si a actiunii radiatiior asupra plantelor,
animalelor si omului, a dus la aparitia radiogeneticii, stiinta care studiaza efectul
radiatiilor asupra ereditatii. Importanta cercetarilor
de radiogenetica este incontestabila, mai ales prin elaborarea unor
metode care asigura protectia organismelor de efectul nociv al
radiatiilor.
Deosebit de
ample au fost investigatiile privind rolul diferitelor componente
biochimice ale celulelor in procesul de transmitere a informatiei
ereditare de la parinti la urmasi. Descifrarea codului genetic a
pus in evidenta legatura dintre acizii nucleici si biosinteza
proteinelor. S-a reusit sa se studieze ereditatea la nivelul
structurilor moleculare si, procesele biochimice si biofizice care au
loc in celula vie. Astfel, a luat nastere genetica moleculara, care
studiaza ereditatea cu ajutorul metodelor moderne de investigatie ale
biofizicii, biochimiei, virusologiei, matematicii, etc.
Cunostintele
actuale asupra mecanismelor ereditare, asa cum actioneaza ele la
nivelul individului, desi departe de a fi complete, au devenit foarte
specializate si de o mare subtilitate.
In
acelasi timp, s-a recunoscut ca exista forme de organizare
superioare nivelului individual, de importanta hotaratoare in
natura. Acestea reprezinta grupari de indivizi ai aceleeasi
specii ce interactioneaza unii cu altii, alcatuind
populatii si grupari de populatii ale diferitelor specii,
aflate in interactiune, care alcatuiesc comunitatile
biotice. Pentru a intelege complexitatea interactiunilor la nivelele
superioare celui individual, trebuie abordate problemele populatiei. De
aceasta latura a stiintei se ocupa genetica populatiilor.
Genetica, ca
si alte discipline, a cunoscut momente de incertitudine, cand i-au fost
puse la indoiala teoriile de baza. Astfel este momentul Temin, care
descopera enzima reverstranscriptaza,
capabila sa realizeze sinteza de ADN pe matrita ARN si
demonstreaza ca moleculele dezoxiribonucleice nu sunt singurele purtatoare
de informatie ereditara.
Un alt moment
este acela al descoperirii prionilor,
particule infectioase responsabile de asa-numitele encefalopatii subacute spongioase
transmisibile (de exemplu, bolile scrapie
si Creutzfeldt-Jacob). Stanley
Prusiner a reusit sa demonstreze ca agentul cauzal al acestor
maladii nu este altceva decat o molecula proteica constituita
din 254 de reziduuri aminoacidice, ea fiind varianta modificata a unei
proteine ce exista in mod obisnuit in celulele nervoase. Proteina
normala, produs al unei gene numite gena
prionilor, se deosebeste de cea patologica doar prin
conformatia tridimensionala, ambele avand aceasi
conformatie aminoacidica. Esential este faptul ca proteina
prionoca modificata se comporta ca un veritabil agent patogen,
transmitand boala la indivizii sanatosi, cu conditia
ca acestia sa posede in celule o copie normala a genei
prionilor. Printr-un mecanism inca neelucidat, proteina
infectioasa, ajunsa intr-un asemenea organism, transforma
prionii normali in prioni patologici. De aceea a fost avansata ipoteza
ca proteina prionica scrapie detine informatia
necesara acestei transformari. Daca cercetarile ulterioare
o vor confirma, ipoteza lui Prusiner, atat de contestata in lumea
stiintifica mondiala, va trebui sa fie acceptata
macar ca o exceptie de la dogma centrala a geneticii.
*
Doresc a-i
multumi pe aceasta cale domnului profesor dr. Ion Bara,
coordonatorul stiintific al acestei lucrari, pentru sprijinul
acordat de-a lungul celor patru ani de facultate, pentru bibliografia pusa
la dispozitie, pentru accesul la Laboratorul de Genetica al
Facultatii de Biologie, dar nu in ultimul rand pentru sfaturile sale
nepretuite care m-au apropiat de persoana dumnealui si de disciplina
pe care o conduce.
I. ISTORIC
I.1. Din istoricul cercetarilor
I.1.1Din istoricul cercetarilor pe
plan mondial
Prima
persoana care a crescut indivizi de Drosophila
melanogaster pe medii nutritive, in laborator, a fost C. W. Woodworth.
Avantajele folosirii indivizilor acestei specii, ca obiect de studiu, au fost
sesizate de catre W. E. Castle, care a si efectuat, de altfel,
primele cercetari pe Drosophila
melanogaster, intre anii 1901 - 1905. Ulterior, el a recomandat
aceasta specie lui T. H. Morgan, care a inceput sa o cultive din anul
1909. Laboratorul din Universitatea Columbia, celebra "Fly Room", a
functionat sub conducerea lui T. H. Morgan intre anii 1910 - 1926.
Impreuna cu ilustrii sai colaboratori (Alfred Henry Sturtevant,
Calvin B. Bridges si Hermann Joseph Müller), Morgan a elaborat istorica teorie cromosomiala a
ereditatii (1910 - 1915), prezenta in lucrarea Mechanisms of Mendelian Inharitance
(publicata in anul 1915). Morgan si colaboratorii sai au
demonstrat, experimental, legatura dintre principiile stabilite de Mendel
pentru mostenirea caracterelor si cromosomi, aratand ca
genele, purtatori materiali ai informatiei ereditare, sunt localizate
pe cromosomi, intr-o ordine liniara. Mai mult, ei au demonstrat ca
genele plasate pe acelasi cromosom au tendinta de a se transmite in
bloc, in succesiunea de generatii. Fenomenul a fost denumit, de Morgan
si scoala creata de el, "linkage"
(inlantuire).
Astfel, Drosophila melanogaster, avand patru
perechi de cromosomi, are patru grupe de inlatuire. Pe baza
experimentelor de incrucisare, s-a putut determina carui grup de
linkage (I, II, III sau IV) ii apartine o gena, precum si
pozitia pe care aceasta o ocupa in cadrul cromosomului respectiv.
Apoi s-a dovedit
ca la femelele de Drosophila
melanogaster linkage-ul nu este complet, in sensul ca uneori are loc
schimbul de gene intre cromosomii omologi, fenomen denumit de T. Morgan "crossing-over". Prin
investigatiile minutioase si repetate s-a demonstrat ca frecventa
crossing-over-ului este dependenta de distanta dintre genele din
acelasi grup de linkage. Cu cat distanta dintre gene este mai mare,
cu atat creste frecventa recombinarilor. Pe baza acestor
observatii, A. H. Sturtevant (unul din discipolii lui Morgan) a realizat
prima harta cromosomiala, luand ca unitate de distanta
asa-numita unitate Morgan,
echivalenta cu un procent de recombinare.
Mai tarziu,
in 1920, T. H. Morgan s-a mutat la Institutul de Tehnologie din California,
impreuna cu Bridges si Sturtevant. Tot in 1920, H. J. Müller (un alt
discipol al lui Morgan) s-a mutat la Universiatatea din Austin, unde a
colaborat cu J.T. Petterson, T. S. Painter si W. S. Stone. Experimentele
de mutageneza efectuate de Müller asupra Drosophilei au demonstrat ca radiatiile Röentgen
provoaca mutatii (la indivizii acestei specii), infirmand
conceptia autogenetica, potrivit careia factorii exteriori nu
pot modifica ereditatea organismelor.
Putin
mai tarziu, in Rusia, Nicolai Petrovici Dubinin si colaboratorii sai
au identificat o serie intreaga de mutatii noi.
In anul 1928
Hanson si Heys provoaca aparitia de mutatii la Drosophila melanogaster cu ajutorul
radioului. Ulterior Auerbach (1949) publica o cronica detaliata
a mutatiilor determinate de factori chimici, asemanatoare celor
produse de razele X.
In paralel cu
experimentele de hibridare si mutageneza chimica, au fost
efectuate si investigatii citologice asupra musculitei de
otet. Astfel, in 1908, Stevens stabileste pattern-ul cromosomial al
femelei, dar nu si pe cel al masculului. Bridges (1914), identifica
cromosomul Y la o femela care
prezenta formula 6A + XXY. Metz
evidentiase cromosomii drosofilei, tot in 1914, identificand cele patru
perechi de cromosomi ale femelei, dar cromosomul Y de la mascul ramasese neobservat. Abia in 1935, Darlington a demonstrat ca
linkage-ul heterosomal complet si indestructibil, la masculii de Drosophila melanogaster, se datoreaza faptului
ca heterosomii X si Y nu sunt omologi.
Studiile
biochimice efectuate de catre Buternandt si colaboratorii sai au
constatat ca gena v+(v = vermilion, ochi de culoare
rosu aprins) controleaza conversia triptofanului in kinurenina.
Mai tarziu s-a demonstrat ca un al doilea pas pe calea sintezei
pigmentului maro din ochii de Drosophila
melanogaster este controlat de gena cn+ (cn = cinnabar, ochi
purpurii), a carei mutatie blocheaza conversia kinureninei in
3-hidroxikinurenina. Aceste cercetari biochimice au dat nastere
conceptiei "o gena - o enzima", care ulterior a fost reformulata.
Cercetarile
efectuate de catre Edward B. Lewis, la inceputul anilor 1940, au condus la
elaborarea asa-numitului test cis-trans
pentru demonstrarea alelismului de functie la Drosophila melanogaster. Cu ajutorul acestui test se poate afla
daca doua mutatii ce afecteaza un anumit caracter, sunt
localizate in aceeasi gena sau in gene diferite. Testul trans, numit si test de
complementare, constituie, actualmente, o modalitate de definire a genei - unitatea fundamentala de
nefunctie a materialului ereditar.
Conceptia
clasica despre gena ca unitate de functie, de mutatie
si de recombinare, a suferit modificari esentiale in urma
studiilor lui N. P. Dubinin asupra mutatiilor scute (sc) la Drosophila.
Analizand raporturile de alelism dintre scute-1, scute-2, scute-3, Dubinin a
ajuns la concluzia ca gena nu se comporta ca o entitate
indivizibila in procesul de mutatie (asa cum postulase T.
Morgan). In anii 1929 - 1933, pe baza observatiilor asupra
comportamentului alelelor scute (14 alele notate sc1-sc14), N. P.
Dubinina elaborat teoria divizibilitatii genei, care a devenit mai
tarziu un principiu central al geneticii moleculare.
Cercetarile
efectuate asupra Drosophila melanogaster
au contribuit la elucidarea a numeroase fenomene de nivel individual si
populational. Actualmente, studiile asupra musculitei de otet se
inscriu in sfera geneticii
dezvoltarii, ramura care
incearca sa elucideze mecanismele complexe ale reglajului genetic la
eucariote, mecanisme direct implicate in procesele de diferentiere si
metamorfoza.
De la
infiintarea celebrului laborator al lui T. H. Morgan, numarul celora
care studiaza drosophila a crescut necontenit. Sute de laboratoare
investigheaza astazi genele si cromosomii acestei miraculoase
vietati. In Statele Unite ale Americii exista doua mari
centre care mentin culturi de mutante si anume: Drosophila Stock Center de la Institutul de tehnologie din
California (Pasadena), si Mid-America Stock Center de la Universitatea de
Stat din Bowling Green.
Pe internet a
fost creata baza de date Flybase, care poate fi accesata pe sit-urile
Gopher sau World Wide Web (https://morgan.harvard.edu).
Aici sunt prezentate 23878 de alele pentru 8676 de gene din genomul Drosophilei melanogaster.
I.1.2. Din istoricul cercetarilor efectuate in Romania
La
initiativa lui G. Marinescu ia nastere in anul 1936 Societatea de Genetica din Romania
care a contribuit la dezvoltarea cercetarilor de genetica si la
schimbul de idei intre oamenii de stiinta din acest domeniu.
In 1936 Gh.
Stroescu isi trece la Sorbona teza de doctorat in stiinte, sub
coordonarea profesorului N. Caullrey, cu un subiect de genetica
experimentala la Drosophila
melanogaster, tratand o noua serie de alele multiple. Lucrarea avand
titlul "Une nouvelle serie d alleles multiples determinant le caractere-scalloped et
localisee dans le cromosome sexuel, chez Drosophila melanogaster
Tot in anul
1936 a avut loc prima comunicare, din Romania, privind mutantele de Drosophila melanogaster, lucrarea fiind
elaborata de G. K. Constantinescu si Veturia Derlogea.
Prin
atasatul agricol al S.U.A. in Bucuresti, profesorul G. K.
Constantinescu a solicitat in 1941 Departamentului de Genetica de la
Carnegie Institute of Washington, suse de Drosophila melanogaster cu mutatii pe anumiti cromosomi,
fapt ce a permis ulterior o analiza sistematica a mutatiilor ce
se obtineau in laborator, cu precizarea locusului in care se aflau.
La
intoarcerea din Gemania, unde a fost trimis pentru specializare, Gh. Radu a
adus numeroase carti de genetica si toate mutantele de Drosophila, in numar de 400, pe
care le avea profesorul Timofeev-Resovsky (stocul de mutante din Berlin, la
epoca respectiva, fiind cel mai valoros si mai complet din Europa).
Primele
cercetari genetice pe Drosophila
melanogaster, realizate de un roman, au fost acelea efectuate de Gheorghe
Radu in Germania in 1943. Intr-o suita de sapte lucrari
experimentale a studiat cu ajutorul razelor X inducerea mutatiilor in
gametii de Drosophila. Utilizand
diferite doze de radiatii Röentgen, a evidentiat, indeosebi, rata
translocatiilor intre cromosomii perechilor II si III.
Toate cele
sapte lucrari au fost publicate in revista "Naturwissenschaft".
Gh. Radu in
colaborare cu Kanelli continua seria de studii abordand, in cadrul a trei
lucrari, studiul citogenetic al cromosomilor perechii I la Drosophila melanogaster prin iradierea
masculilor cu raze X, studiul comparativ al translocatiilor genetice intre
cromosomii perechilor II si III,
precum si studiul altor aberatii cromosomiale precum inversiunile,
duplicatiile si gapsurile.
Rezultatele
cercetarilor analizei genetice asupra populatiei de Drosophila melanogaster din
Bucuresti, identifica mutatiile: forked, abnormal venation
si white. S-au identificat la Drosophila melanogaster sapte
mutatii, dintre care patru localizate in cromosomii perechii I si
trei in cromosomii perechii II.
Dupa cel
de-al doilea razboi mondial, cercetarile de genetica,
teoretica si experimentala, s-au amplificat si
diversificat, infiintandu-se catedre si laboratoare de cercetare in
diferite institutii stiintifice.
In anul 1964
s-a tinut la Universitatea din Bucuresti primul simpozion
national de genetica, la care s-au prezentat peste 70 de referate
stiintifice si comunicari. Al doilea simpozion de
genetica, desfasurat in 1967 la Bucuresti, a avut printre
participanti cunoscuti specialisti straini. In cadrul
acestui simpozion, Gh. Radu prezinta comunicarea "Modificarile
citogenetice in cromosomii de Drosophila
melanogaster prin iradieri fractionate cu raze X".
La Facultatea
de Biologie din Iasi, introducerea studiilor asupra drosofilei este
asociata cu numele Prof. Dr. Iordachi Gh. Tudose (1937 - 1997).
Dintre
lucrarile sale amintim: Modificari
in desfasurarea ciclului vital la Drosophila melanogaster sub
influenta efedrinei si actiunea acestei substante asupra
cromosomilor uriasi, 1969, in colaborare cu R. Brandsch (actualmente
profesor la Universitatea "Albert Ludwigs" din Freiburg, Germania); Influenta acidului nicotinic asupra
desfasurarii ciclului vital la Drosophila melanogaster,
1977, in colaborare cu Alexandra Rabacov.
In revistele
de specialitate din Romania sunt publicate numeroase articole care au ca obiect
de studiu Drosophila melanogaster.
S-au realizat numeroase experiente privind organizarea cromosomilor
uriasi, actiunea selectiei asupra diferitelor populatii de Drosophila, fenomenul de curtare
realizat de masculi, obtinerea de mutante sub actiunea
substantelor chimice si agentilor mutageni fizici, fenomenul de
apoptoza si rolul mortii celulare in dezvoltarea si
metamorfoza Drosophilei.
II. DROSOPHILA
MELANOGASTER - CARACTERIZARE GENERAL-BIOLOGICA SI ECOLOGICA
Aceasta
specie a fost descrisa la mijlocul secolului al XIX- lea sub numele de Drosophila ampelophila, ceea ce
inseamna "iubitoare de vita de vie" (gr. ampelos =
vita de vie, philos = iubitor ). Ulterior i s-a dat numele de Drosophila melanogaster, adica
"iubitoare de roua, cu abdomenul negru" (gr. drosos = roua, apa,
lichid; philos = iubitor; melas = negru; gastris = stomac).
II.1. Incadrarea sistematica a speciei Drosophila melanogaster
Specia Drosophila melanogaster apartine
genului Drosophila din Familia Drosophilidae, Grupul Acalyptratae, Sectiunea Schizophora, Subordinul Cyclorrhapha; Ordinul Diptera, Diviziunea Endopterigota (Holometabola),
Subclasa Ptrerygota, Clasa Insecta, (Hexapoda), Increngatura Artropoda.
Genul Drosophila este impartit
intr-un numar de subgenuri, si anume: Hirtodrosophila, Pholadoris,
Dorsilopha, Phloridorsa, Sophophora
si Drosophila. Unele dintre
subgenuri au fost divizate in "grupuri de specii". De exemplu, subgenul Sophophora cuprinde sapte grupuri
de specii, dintre care Saltans, Willistoni si Melanogaster, ultimului apartinandu-i Drosophila melanogaster.
Numarul
speciilor din genul Drosophila este
de aproximativ 800, majoritatea caracterizate prin polimorfism cromosomial.
O
problema disputata este incadrarea Familiei Drosophilidae intr-un anumit subordin. Astfel, unii autori
plaseaza aceasta familie in Subordinul Brachycera, considerand nepotrivita crearea unui nou subordin
- Cyclorrapha - in cadrul Ordinului Diptera. Diferenta majora
dintre cele doua subordine amintite (invocata de sistematicienii ce
recunosc Subordinul Cyclorrapha) este
forma deschiderii prin care adultul emerge din puparium, si anume: forma
literei T, la Brachycera si forma
circulara la Cyclorrapha.
Mentionam
ca denumirea de Acalyptratae
semnifica prezenta caliptrelor foarte mici la idivizii ce fac parte
din acest grup, iar cea de Schizophora,
prezenta scizurii frontale de forma literei U.
II.2. Date morfologice
Drosophila melanogaster are aproximativ 2 mm lungime,
avand corpul moale de culoare gri-galbuie si doi ochi compusi
mari, de culoare rosie-caramizie (la tipul salbatic). In
regiunea capului mai prezinta trei ochi simpli numiti oceli si
doua antene mici, aristate, formate din cate trei segmente; pe cel de-al
treilea segment antenal se afla arista. Aparatul bucal este adaptat pentru
lins si supt, drosofila consumand hrana lichida sau lichefiata
in prealabil cu ajutorul secretiei glandelor salivare.
Ca toate
dipterele, musculita de otet prezinta doua aripi mari,
mezotoracice, aripile de pe metatorace fiind transformate in haltere (organe de
echilibru). Pe fiecare segment toracic se afla cate o pereche de picioare
adaptate la mers.
Numarul
segmentelor abdominale vizibile este diferit la cele doua sexe -
sapte la femela si cinci-sase la mascul. Acest numar
mic de segmente la masculi este consecinta fenomenului de telescopare a
ultimelor segmente abdominale.
Adultii
de Drosophila melanogaster
prezinta dimorfism sexual, manifestat atat prin diferente
dimensionale, cat si morfologice. Femelele sunt mai mari decat masculii.
Au abdomenul oval cu extremitatea posterioara putin
ascutita. La masculi varful abdomenului este rotunjit si colorat
in maro-negru.
Numarul
de omatidii ce intra in alcatuirea ochiului complex este diferit la
cele doua sexe, si anume: 780 la femela, respectiv, 740 la
mascul.
Masculii
prezinta asa numitul "pieptene sexual" care serveste la
imobilizarea femelei in timpul acuplarii. Acesta este format din perisori
negri, rigizi, ce se gasesc pe primul articol tarsal al picioarelor
anterioare (perechea I), mai precis, in apropierea articulatiei dintre
tibie si tars. Evident ca diferentele cele mai insemnate dintre
mascul si femela tin de organizarea interna, mai ales de
organizarea sistemului reproducator.
Sexul
larvelor se determina tinand cont de pozitia si
marimea gonadelor. Acestea sunt localizate in partea antero-laterala
a larvei, in corpii grasi. Testicolele sunt mult mai mari decat ovarele
(de trei ori mai mari) si au o forma ovala. Ele pot fi observate
cu ochiul liber datorita transparentei tegumentului larvei.
II.3. Ciclul de viata la Drosophila melanogaster
Ca toate
dipterele, Drosophila melanogaster prezinta
metamorfoza completa (este o insecta holometabolica),
ciclul sau vital parcurgand stadiile de ou, larva, pupa si
adult. Durata ontogeniei variaza in functie de temperatura,
umiditate, compozitia mediului nutritiv etc.
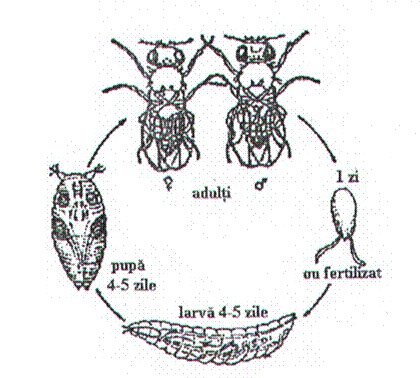
Figura 1. Desfasurarea ciclului vital la Drosophila melanogaster (Autorul figurii necunoscut)

Figura 2. Stadiul de ou
(Autorul
necunoscut)
Stadiul de ou. Este cuprins intre momentul depunerii oului de catre
femela adulta si cel al aparitiei larvei corespunzatoare.
Oul este de tip centrolecit, are o lungime de 0,5 mm si forma ovala,
cu partea dorsala mai bombata decat cea ventrala,
aplatizata. Membrana externa, numita si chorion, este
groasa, opaca, si prezinta un model de semne hexagonale. In
partea anterioara si dorsala oul prezinta o pereche de
filamente
(apofize), acestora fiindu-le
atribuit rolul de a impiedica scufundarea sa in mediul lichid.

Figura 3. Stadiul de
larva (Autorul necunoscut)
Stadiile larvare la Drosophila melanogaster sunt in numar de trei, fiind separate
intre ele prin doua naparliri.
Larva de stadiu I paraseste
oul prin micropil, este mica, foarte mobila si se
hraneste cu mare repeziciune. Este o larva de tip apod
(fara apendici), acefala (capsula cefalica este
infundata comlet in torace). Aparatul bucal de tip primitiv (adaptat
pentru rupt si mestecat), prezinta mandibule puternice, ca niste
carlige, cu ajutorul carora larva patrunde in materialul nutritiv.
Acest prim stadiu larvar dureaza aproximativ 24 de ore.

Figura 4.
Stadiul de pupa (Autorul necunoscut)
Pe
masura ce larva creste, cuticula sa chitinoasa devine
neancapatoare, astfel, ca larva o paraseste
si isi sintetizeaza o noua cuticula
(naparleste).
Larva de
stadiul II apare in urma primei
naparliri. Aceasta este mai mare si continua sa se
hraneasca timp de 24 de ore, dupa care sufera inca o
naparlire.
Larva de
stdiul III apare dupa cea de-a doua naparlire (dupa aproximativ
sase zile de la fertilizare). Aceasta poate atinge 4,5 - 5 mm lungime,
dezvoltandu-se pe parcursul a 48 de ore.
Stadiul de
pupa. Are o durata variabila, acest parametru fiind profund
influentatde temperatura. La Drosophila
melanogaster pupa este de tip coarctat "pupa butoias", o
varianta a pupei adectice (cu mandibule nesclerificate) de tip liber,
caracterizata prin aceea ca larva de stadiul III se impupeaza in
interiorul ultimei exuvii larvare. La inceput, invelisul pupal chitinos
este alb si moale. Odata cu scurtarea corpului larvei, el se
intareste si se inchide la culoare, fiind numit si
puparium. Are forma ovala, masoara 2,5 - 3 mm si
prezinta in partea anterioara doua prelungiri ce corespund
spiraculelor evaginate ale fostei larve.
Stadiul de imago. In urma transformarilor ce
au loc in interiorul invelisului pupal, se formeaza adultul, care
eclozeaza printr-o deschidere circulara din regiunea anterioara
a pupariumului.

Figura 5. Pereche de
adulti apartinand speciei Drosophila
melanogaster (Autorul necunoscut)
La inceput
insecta este foarte lunga, cu aripile nedesfacute, lipite de corp,
si are o culoare deschisa. Aceste caracteristici ale adultilor
nou eclozati constituie criteriile care ii deosebesc de insectele
batrane. Dupa 24 de ore de la iesirea din pupa indivizii
ating maturitatea de reproducere. Astfel femelele vor accepta un mascul pentru
a fi fertilizate. Interesant, s-a constatat ca femelele de Drosophila melanogaster sunt monogame.
Desfasurarea ciclului
vital la Drosophila melanogaster.
Inainte de a
prezenta etapele dezvoltarii la Drosophila
melanogaster, consideram necesara definirea altor doua
procese - determinarea si diferentierea.
Determinarea reprezinta evenimentul reglator ce stabileste un
pattern (model) specific al activitatii genelor, caracteristic pentru
un tip dat de celula. Cu alte cuvinte, determinarea corespunde unei
programari, programare anterioara specializarii.
Diferentierea (corolar al diferentierii)
constituie conditia adoptata de celulele specializate structural
si functional.
Procesul
complex al dezvoltarii la musculita de otet parcurge cateva
etape esentiale, si anume: embriogeneza, cresterea larvara,
impuparea si metamorfoza.
Dezvoltarea embrionara are loc consecutiv
fecundatiei si se desfasoara in interiorul membranei
oului. Oul nefecundat, format in ovarul femelei, este expulzat in uter, unde
are loc fecundatia.
Stadiul
initial al dezvoltarii embrionare consta in formarea primilor
doi nuclei prin diviziunea mitotica a nucleului zigotic, aceasta avand loc
imediat dupa fertilizare. Timpul necesar pentru un ciclu de replicare a
ADN si o mitoza este de aproximativ 10 min. Dupa doua
diviziuni sincrone, cei 512 nuclei rezultati migreaza spre
suprafata externa a oului (cortex), formand asa numitul
blastoderm sincitial. Aproximativ 40 de nuclei migreaza, in schimb,
la polul posterior al oului, unde vor da nastere celulelor polare. Nucleii
din cortex continua sa se divida, se inconjoara cu
citoplasma si se inchid in membrana, formand ceea ce se
numeste blastoderm celular. Acest stadiu apare aproximativ la 2,5 ore de
la fertilizare si se prezinta sub forma unui singur strat de celule,
situat la exteriorul embrionului. Trebuie precizat ca dezvoltarea
embrionara incepe cu stabilirea polaritatii oului, de-a lungul
axelor antero-posterioara si dorso-ventrala. Acest proces
prezinta importanta majora, fiind demonstrat faptul
ca, mutatiile cre afecteaza polaritatea sunt cauza
dezvoltarii structurilor anterioare in regiunea posterioara,
respectiv a celor doua dorsale in partea ventrala a oului.
In stadiul de
blastoderm, soarta fiecarei regiuni a oului este deja stabilita.
Cercetarile au dovedit ca trasaturile majore ale planului
corpului larvei si adultului, sunt prezente inca din acest stadiu
timpuriu al embriogenezei. Proiectiile bidimensionale ale organelor
externe ale adultului pe suprafata blastodermului, sunt cunoscute sub
numele de "fate maps" - harti ale destinului.
Celulele
blastodermului urmeaza doua cai posibile de dezvoltare. Astfel,
majoritatea lor vor continua sa se divida, se vor diferentia
si vor asigura formarea embrionului. O parte dintre blastomere se vor
separa de celelalte, ele constituind celulele de origine ale tesuturilor
viitorului adult. Aceste celule programate sa genereze structuri adulte se
vor organiza in asa numitele discuri imaginale. Exista 12 perechi de
discuri bilaterale si un singur disc genital, de exemplu: o pereche pentru
ochi si antene, trei perechi pentru picioare, o pereche pentru aripi, o
pereche pentru haltere, etc.
Alte celule
ale blastodermului vor forma structurile interne si externe ale larvei.
Desi planul viitorului organism adult este prezent inca din stadiul
de blastoderm, segmentele acestuia nu se disting. Segmentele sunt structuri
morfologice vizibile, atat la adult, cat si la larva.
Embrionul de
10 ore prezinta trei segmente toracice si opt abdominale, separate
prin santuri, corespunzand segmentelor viitorului adult. In acest
stadiu al dezvoltarii, celulele fiecarui segment sunt determinate
sa formeze fie un compartiment anterior (jumatatea anterioara a
respectivului segment), fie un compartiment posterior (jumatatea
posterioara a acestuia).
Segmentul nu
este altceva decat unitatea de structura a embrionului, in timp ce
compartimentul constituie unitatea de dezvoltare formate din celulele
viitoarelor discuri imaginale. Descendentii acestor celule, numite si
celule fondatoare, nu pot trece dintr-un compartiment in altul, astfel ca
la embrionul tardiv si la adult, compartimentul se distige prin aceea
ca include numai celulele provenind din linia fondatoare
corespunzatoare.
Trebuie
precizat ca in stadiul de blastoderm, cea mai mare parte a embrionului
este divizata intr-o serie de compartimente alternative, anterioare
si posterioare. Un segment adult insa, consta dintr-un singur compartiment anterior, urmat de unul
posterior.
La nivelul
blastodermului a fost sugerat si un
alt tip de segmentare, situatie in care santurile de separare
sunt vizibile intre asa numitele parasegmente. Un parasegment consta
din compartimentul posterior al unui segment si compartimentul anterior al
segmentului urmator. Astfel, granitele dintre segmente corespunde cu
centrele parasegmentelor.
Toate aceste
procese sunt controlate de genele de segmentare, ale caror mutatii
afecteaza atat numarul cat si polaritatea segmentelor corpului.
Regiune
posterioara a oului contine componente citoplasmatice numite granule
polare, iar nucleii care migreaza in aceasta regiune formeaza
asa numitele celule polare, care sunt precursorii celulelor germinale ale
adultului. Daca regiunea posterioara a oului este distrusa inainte
de migrarea nucleilor, sau daca aceasta este impiedicata prin
ligatura embrionului, musculita adulta se dezvolta, totusi,
dar este sterila, neavand celule germinative. Transplantul de
citoplasma polara in regiunile anterioare ale oului, induce formarea
celulelor polare in regiuni neobisnuite. Asemenea celule polare induse pot
genera celule germinale functionale daca sunt inlaturate
chirurgical si injectate in regiunea posterioara a embrionilor
marcati genetic. Astfel, citoplasma specifica de la polul posterior
al oului, propaga in nucleii ce migreaza la acest pol, o stare de
determinare care, conduce la o forma specifica de diferentiere.
A fost
formulat postulatul conform caruia intr-un segment sau compartiment
individual, una sau mai multe gene selectoare (detinatoare de
informatie pozitionala) determina o cale particulara
de dezvoltare pentru celulele din acest compartiment.
Mutatiile
acestui sistem de selectie, numite mutatii homeotice, modifica
starea de determinare a celulelor dintr-un segment sau compartiment, astfel
ca ele genereaza structuri intalnite normal in alte segmente. De
exemplu, mutatia "tumouros head" (engl. = cap tumoral) transforma
capul in ultimul segment abdominal si in genitalii.
La
temperatura de 25o C, dezvoltarea embrionara dureaza
aproximativ 22 de ore si se incheie cu eclozarea larvei de stadiul I. Pe
parcursul tuturor celor trei stadii larvare, din momentul aparitiei
primului stadiu si pana la formarea pupariumului, celulele larvare nu
se divid, desi larva creste considerabil.
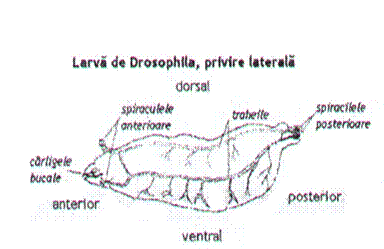
Figura 6.
Larva de Drosophila melanogaster.
Privire laterala. (Autorul necunoscut)
Aceasta crestere se
datoreste exclusiv maririi taliei celulelor, care este
insotita de replicarea ADN si formarea cromosomilor politenici
(politenizare). Spre deosebire de celulele larvare, cele adulte (reprezentate
de celulele discurilor imaginale) se divid in timpul acestei perioade.
Astfel, cresterea
discurilor imaginale are loc pe seama diviziunii celulelor acestora. In timpul
celui de-al doilea stadiu larvar, discul imaginar al aripii este constituit din
50 de celule, la 35 de ore de la fertilizare. Dupa 122 de ore,
numarul celulelor respectivului disc atinge 30000.
La 25oC, primul
stadiu larvar dureaza 24 de ore, astfel ca, prima naparlire are
loc la 46 de ore de la fertilizare. Al doilea stdiu larvar este cuprins intre
cele doua naparliri (cea de-a doua naparlire larvara
survine dupa 70 de ore de la fertilizare) si dureaza alte 24 de
ore. Ultimul stadiu larvar atinge dimensiunea maxima dupa 48 de ore
de la cea de-a doua naparlire.
Urmatoarea etapa a
dezvoltarii la Drosophila
melanogaster o constituie impuparea.
Problema formarii pupei
si cea a pupariumului reprezinta motivul unei dispute indelungate
intre specialistii in fiziologia insectelor. Este evident ca cele
doua procese sunt identice. Astfel:
formarea
pupariumului conduce la aparitia stadiului de prepupa si poate
fi considerata prima parte a unei naparliri, epiderma fiind inca
lipita de prepupariumul format in vechea cuticula a larvei de stadiul
III;
finalizarea
acestei naparliri si desprinderea epidermei fostei larve de puparium
(ultima exuvie larvara) constituie tocmai procesul de impupare.
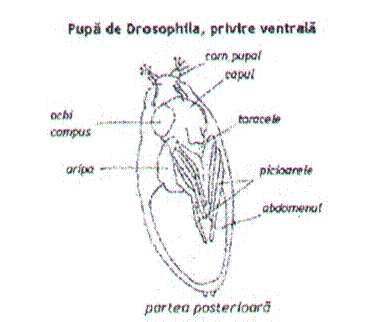
Figura 7. Pupa de Drosophila melanogaster. (Autorul necunoscut)
Formarea
pupariumului, a pupei si metamorfoza au fost studiate de multi
cercetatori, pe parcursul unei lungi perioade. Se cunosc 51 de stadii
vizibile in timpul metamorfozei si descrie 24 de stadii utilizabile in
analiza experimentala.
Cu opt ore
inainte de a incepe formarea prepupariumului, larva de stadiul III nu se mai
hraneste, paraseste mediul nutritiv, in cautarea
unei suprafete relativ uscate. Aceasta etapa se mai numeste
stadiul larvar hoinar. Dupa aproximativ 120 de ore de la fertilizare (la
25oC), incepe procesul de formare a prepupariumului odata cu
aparitia unei prepupe de culoare alba si care prezinta in
partea anterioara spiraculele evertate, ea incetand sa se miste
(timpul zero). Prepupa (si ulterior pupa) ramane fixata pe substarat
prin intermediul unei proteine lipicioase secretata de glandele salivare ale
larvei. In decursul a 20 de minute - o ora, culoarea prepupei devine maro.
Dupa patru ore de la aparitia prepupei, in abdomenul abdomenul
acesteia se formeaza o bula de aer, astfel ca organismul
respectiv capata proprietatea de flotabilitate. Imediat dupa
acest eveniment incepe formarea pupei propriu-zise, stadiu care atinge forma
completa la aproximativ 12 ore de la momentul zero - inceperea formarii
prepupariumului.
Ciclul de
dezvoltare continua cu metamorfoza, etapa in care au loc intense
procese de histoliza si histogeneza. Majoritatea celulelor
larvare sufera autoliza, cu exceptia tuburilor Malpighi si a
unor celule nervoase. Tesuturile adulte iau nastere din celulele discurilor
imaginale. Acestea pot fi grupate in doua categorii, si anume:
discuri ale carei celule au suferit diviziuni in timpul vietii
larvare, dar au incetat sa se divida in timpul impuparii,
si celule imaginale care se divid in perioada timpurie a metamorfozei, cum
sunt histoblastele abdominale.
|
Stadiul de dezvoltare
18,5oC 25oC 29oC
|
|
Aparitia larvei de stadiu I dupa 41 ore dupa 21 ore dupa 20 ore
Formarea prepupariumului dupa 234 ore dupa 118 ore dupa 110 ore
Ecloziunea adultului dupa 440
ore dupa 221 ore dupa 190 ore
|
Tabelul 1. Durata ciclului vital la tipul
salbatic Oregon R, pentru diferite stadii de dezvoltare si la diverse
temperaturi
Celulele
discurilor imaginale cresc, sufera miscari morfogenetice si
se diferentiaza, conducand la constituirea adultului de Drosophila melanogaster. Precizam
faptul ca gonadele adultului se
diferentiaza pornind de la gonadele larvare (care, la randul lor
isi au originea in celulele polare), diviziunea reductionala
si formarea gametilor avand loc in timpul metamorfozei. Astfel,
adultii sunt apti de imperechere, in medie, dupa 220 de ore (la
25oC) de la fertilizare.
II.4. Raspandire si ecologie
II.4.1. Habitatul natural si conditiile de
laborator
Indivizii
speciei de Drosophila melanogaster
sunt frecvent intalniti in natura, pe fructele care au inceput
sa fermenteze (mere, pere, prune, struguri, etc.). vara pot fi
intalniti in vii, gradini, livezi sau in magazii, beciuri cu fructe
si legume (mai ales daca acestea sunt alterate), in depozite, pe
tescovina aruncata si, in general, in locurile insalubre.
Aceasta specie nu prezinta diapauza, iernand in magazii,
depozite, beciuri, etc.
In
laborator, musculita de otet poate fi cultivata pe medii
sintetice, care contin zahar si drojdie - componente ce
mijlocesc procesul de fermentatie. Compozitia mediului are
consecinte asupra ciclului vital, viabilitatii si
prolificitatii insectei.
Unii autori
impart mediile nutritive in cinci categorii, in scopul in care sunt folosite:
mediile standard, utilizate in orice scop
- mentinerea culturilor, pregatirea incrucisarilor, etc;
mediile imbogatite, folosite cu
scopul de a obtine larve mari necesare pentru studiul cromosomilor
politenici si pentru studiile biochimice;
mediile "instant", disponibile in
comert, usor de preparat in caz de urgenta;
mediile axenice, pentru cresterea
musculitelor in conditii sterile;
mediile sintetice definite, pentru
studiile biochimice.
II.4.2. Temperatura - factor fizic ce
influenteaza dezvoltarea Drosophilei
melanogaster
Temperatura,
ca si compozitie a mediului de cultura, influenteaza
puternic desfasurarea ciclului vital la Drosophila melanogaster. La temperaturile mai mari de +31oC,
femela devine, partial sau chiar total sterila. In regiunile cu
umiditate sporita, musculita de otet poate tolera temperaturi
mai ridicate ale aerului.
Temperatura
optima pentru dezvoltarea indivizilor de Drosophila melanogaster este de +24 - +25oC, la
aceasta temperatura ciclul vital dureaza 9 - 10 zile (stadiului
de ou ii revin 24 de ore, celui de larva 4 zile si pupei tot 4 zile).
Astfel, intr-un an se pot obtine 40 de generatii.
La
temperaturi mici, ciclul de dezvoltare al insectei se lungeste. Astfel, la
+ 20oC, ciclul vital se prelungeste pana la 16 zile (larva
8 zile, pupa 6 - 8 zile), pe cand la + 10oC durata ontogeniei ajunge
la 75 de zile (larva de 57 de zile, pupa 13 - 18 zile).
Drosophila melanogaster prezinta mai multe tulpini
"termosensibile". In majoritatea cazurilor temperatura permisiva este de +
18oC sau + 20oC, iar temperatura restrictiva este de
+ 29oC.
II.4.3. Umiditatea - factor fizic ce influenteaza dezvoltarea Drosophilei melanogaster
Umiditatea
optima necesara dezvoltarii normale pentru Drosophila
este de 75 - 85 %. In unele cazuri, acest
parametru poate deveni o problema serioasa.
Un nivel al umiditatii sub 70 % afecteaza profund fertilitatea si viabilitatea
insectelor.
II.4.4. Prolificitatea - factor ce influenteaza dezvoltarea Drosophilei melanogaster
Musculita
de otet prezinta o prolificitate ridicata. In cursul
vietii, o femela depune cateva sute de oua. In majoritatea
cazurilor numarul oualelor depuse este de 200-300. Ameliorarea
conditiilor de viata determina o crestere a
numarului de oua depuse, uneori ajungand pana la 2000. In medie
o pereche de insecte da nastere la aproximativ 170 de
descendenti.
II.4.5. Longevitatea - factor ce influenteaza dezvoltarea Drosophilei melanogaster
Adultii
de Drosophila melanogaster, in
conditii de laborator, traiesc aproximativ 3-4 saptamani.
Longevitatea depinde de conditiile in care sunt crescute insectele,
si anume: temperatura, umiditatea, compozitia mediului nutritiv,
densitatea populatiei, prezenta in mediu a parazitilor, etc.
In
conditii optime, indivizii de Drosophila
pot supravietui chiar 22 de saptamani (153 de zile). Exista
unele mutante care au o viabilitate mult mai mare decat a Wild type (tipul
normal sau salbatic), insa, de obicei, mutantele si liniile sunt
mai putin viabile comparativ cu tipul normal.
II.5. Unii paraziti ai culturilor de Drosophila melanogaster
Se
cunosc trei categorii de paraziti ce afecteaza culturile de Drosophila: fungi, bacterii si
capuse.
Pentru a reduce contaminarea cu fungi se
recomanda folosirea unui mediu axenic, care contine Nipagin M.
Problemele apar in special la "tulpinile" cu crestere lenta, sau in
cazul in care culturile sunt transferate de pe mediul contaminat pe mediul
proaspat. In aceasta situatie, culturile contaminate trebuie
transferate pe mediu continand atat
Nipagen M, cat si acid propionic.
Infectarea cu bacterii este mai putin
frecventa decat contaminarea cu fungi. Totusi, in cazul
infectiilor bacteriene severe, insectele trebuie transferate pe mediu
tratat cu dihidrostroptomicinsulfat si penicilina G.
Cele mai grave contaminari ale
culturilor de Drosophila sunt cauzate
de capuse. Sunt descrise doua clase:
capusele Mesostigmate - se
intalnesc destul de rar, care se hranesc cu oualele de Drosophila si pot compromite cu
usurinta o cultura.
capusele Anoetide - nu
afecteaza musculitele intr-o maniera atat de directa. Ele
se multiplica rapid si se fixeaza pe adulti, mai ales la
nivelul genitaliilor. Indivizii paraziti devin sterili.
Pentru
a preveni infestarea culturilor de Drosophila
cu capuse, noile culturi trebuie tinute in carantina,
examinate cu atentie si tinute cel putin doua
generatii in afara colectiei proprii. Uneori chiar cele mai severe
masuri de carantina sunt insuficiente. In aceste cazuri se
foloseste un acid puternic - Tedion (2,4,5,4
tetracloro-difenilsulfona) sau benzoat de benzil.
II.6. Determinismul sexelor la Drosophila melanogaster
Wilson
E. B. (1905) si Stevens N. M. (1905), citati de Petre Raicu (1980) au
studiat concomitent determinismul cromosomial al sexelor la diferite specii,
si au descoperit ca in acest proces intervin niste cromosomi
specializati, ce au primit denumirea de cromosomi ai sexului sau heterosomi
(gr. heteros = diferit, soma = corp), alosomi
(gr. alos = altul, soma = corp) sau gonosomi (gr. gonos =
samanta genitala, soma = corp). Restul cromosomilor au fost
denumiti autosomi (gr. autos =
el insusi, soma = corp).
La Drosophila melanogaster, 2n = 8, s-a
remarcat ca femelele poseda patru perechi de cromosomi omologi, pe
cand la masculi, sase cromosomi formeaza trei perechi, iar
ceilalti doi cromosomi nu se pot imperechea fiind diferiti atat ca
marime, cat si ca structura. Acesti doi cromosomi
diferiti au fost notati cu X
si Y, ei fiind heterosomii, iar
ceilalti sase sunt cromosomii somatici (autosomii). Pe de alta
parte, femelele au doi heterosomi identici, notati XX. Astfel, femelele sunt homogametice (gr. homoios =
asemanator, gamos = unire, casatorie) producand un singur
tip de gameti, in timp ce masculii sunt heterogametici (gr. heteros =
diferit, gamos = unire, casatorie), producand doua tipuri de
gameti: jumatate au trei autosomi si heterosomul X, cealalta jumatate prezinta trei autosomi si
heterosomul Y. Prin combinarea
probabilistica in procesul fecundatiei a gametilor femeli de un
singur tip cu gameti masculi de doua tipuri, rezulta 50% femele
(XX) si 50% masculi (XY), sex ratio 1:1.
Acest
tip de determinism sexual caracterizat prin aceea ca femelele sunt
homogametice (XX), iar masculii,
heterogametici (XY), a primit
denumirea de tipul Drosophila.
III. ELEMENTE DE GENETICA A
POPULATIILOR SI EVOLUTIE
III.1. Definitia si
caracterizarea populatiei
Populatia, ca entitate reala a lumii vii, poate fi
definita si caracterizata, printre altele, din cel putin
doua puncte de vedere - ca etapa in integrarea sistemelor biotice,
si ca entitate taxonomica, subunitate a speciei.
Specia, ca sistem supraindividual de organizare si
integrare a materiei vii se caracterizeaza, printre altele, printr-o mare
heterogenitate interna, prin diversitate determinata de
insasi structura si functiile elementelor sale componente.
Specia, reprezinta, pe de alta parte, punctul nodal, entitatea care
incheie sirul suprapunerilor unitatilor taxonomice cu a celor
sistemice. In timp ce in ierarhia sistemica specia este integrata in
biocenoza, in ierarhia sistenatica specia este subordonata
genului, apoi familiei, clasei etc. Specia, ca etapa din organizarea
si integrarea materiei vii, desi universala, se
caracterizeaza prin variatie si neechivalenta, in sensul
ca nu poate fi pus semnul egalitatii intre o specie
vegetala si una animala, intr-o specie de la baza arborelui filogenetic, si una de la partea
sa superioara etc. In consecinta pentru a caracteriza
fara echivoc fenomenul specie, se impune gasirea unor criterii
generale dar si universale, proprii tuturor speciilor. Apreciem ca
cea mai completa lista, in acest sens, a oferit-o K. M. Zavadskii, in
lucrarea "Specia si speciatia" (Vid
i vidoobrazovannie - 1968). Cu unele mici modificari, pe care le-am
operat noi, respectivele caracteristici sunt:
Numarul indivizilor. Specia, sistem suparindividual,
se compune dintr-un numar optim de indivizi, care sa-i asigure atat
perpetuarea, cat si integralitatea si teritorialitatea. Nivelul
numeric (autoreglabil) al speciei, reprezinta una dintre
trasaturile calitative ale acesteia.
Tipul de organizare. Specia se caracterizeaza
printr-un anumit genofond, deci printr-o baza ereditara unica
si are o organizare specifica, fapt ce-i asigura caracterul de
unitate aparte in structura si organizarea lumii vii.
Reproducerea. Desi cu un anumit grad de variabilitate
interna, specia are capacitatea de a se perpetua ca unitate aparte,
discreta a lumii vii. Prin acest tip de perpetuare, specia isi
conserva determinismul calitativ.
Discontinuitatea. Specia reprezinta o entitate
biologica diferentiata, discreta.
Determinismul ecologic. Specia este adaptata la
conditii determinate de existenta si este apta de
concurenta. Ea ocupa un loc aparte in circuitul material si
energetic al ecosistemului.
Determinismul geografic. Specia este raspandita
prin indivizii sai, pe un teritoriu strict determinat. Arealul este
considerat drept una dintre caracteristicile esentiale ale speciei.
Diversitatea formelor. Specia este
diferentiata si poseda o anumita structura
interna. Cuprinzand numeroase forme heterogene, specia are integraliatate
si reprezinta un sistem.
Caracterul istoric. Specia are un trecut (actuala
specie provine dintr-o specie ancestrala) si, evident, un viitor.
Specia, deci, nu este un rezultat al evolutiei, ci si un
purtator al acesteia (adica este o formatiune in cadrul
careia se deruleaza procesul evolutiei).
Stabilitatea. Specia are capacitatea de a-si mentine
identitatea (caracteristicile genetice, spatiale etc.) pe o perioada
geologica data.
Integralitatea. Specia nu este o simpla suma de indivizi ci o
comunitate cu un anumit grad de integralitate, asigurat prin interactiuni
intre indivizii ei componenti.
Asa cum
precizam anterior, desi caracteristicile enumerate sunt proprii tuturor
speciilor, nu inseamna ca ele se manifesta la fel la toate
speciile. Pe de alta parte, orice modificare survenita pentru una
dintre caracteristici antreneaza, automat, modificarea tuturor celorlalte
caracteristici, intr-o masura mai mare sau mai mica. Sunt
autori, de pilda, care apreciaza ca la speciile apomictice
integralitatea este mult diminuata, in comparatie cu speciile
amfimitice. De asemnea, caracterul de discontinuitate este considerat ca
diminuat la speciile tinere, in curs de formare si stabilizare. Pe de
alta parte, la speciile vechi (relicte chiar) discontinuitatea este
neta, bine exprimata, dar diversitatea interna poate fi foarte
slaba, mult diminuata. De pilda, la eucalipt, salcie, plop,
rogoz, papadie etc., discontinuitatea speciilor (in cadrul genului)
este foarte slab exprimata.
Iata de
ce , a da o definitie categoriei de specie este dificil, daca nu
chiar imposibil. Sunt nenumarate tentativele de definire a fenomenului
specie dar, deocamdata, nici una nu este suficient de scurta,
explicita si universal acceptata. In majoritatea
definitiilor nu se tine seama de esenta fenomenului specie ci,
mai degraba, de caracterele ei exterioare, formulandu-se "instructiuni"
utile in recunoasterea si delimitarea speciilor de catre
specialisti, in activitatea lor practica. Iata, pe scurt, cum au
evoluat conceptiile, si, implicit, definitiile date speciei.
John Ray
(1686 - 1704) concepea specia ca cea mai mica unitate de oganisme identice
din punct de vedere morfologic, ce se reproduc intre ele si dau
urmasi care le conserva identitatea (asemanarea). Ray
evidentiaza unele caracteristici ale speciei, si anume:
caracterul de grup, capacitatea de inmultire de sine statatoare
a grupului, asemanarea morfologica si fiziologica a
indivizilor. Ray aprecia ca variatiile individuale sunt abateri
intamplatoare de la tipul speciei, abateri care nu depasesc
limitele speciei.
Carl Lineé
(1707 - 1778), pe numele sau adevarat Ingemarsson, considera ca
fenomenul specie are o raspandire generala, ca este unitatea
fundamentala a clasificarii si ca este un fenomen al
naturii, nu un produs al activitatii constiintei. Incercand
o definitie, Lineé aprecia ca specia reprezinta o multitudine de
organisme inrudite, asemanatoare ca structura si care, prin
inmultire, reproduc mereu indivizi asemanatori lor. Spre finalul
lucrarii sale (Systema naturae),
publicata in 1735, pe numai 12 pagini, supraevaluand caracterul
stabilitate, afirma: "Sunt atatea specii
cate forme diferite au fost create la inceput".
Alphonse de
Candolle, in 1819, aprecia ca specia reprezinta: "o totalitate de indivizi,
asemanatori intre ei mai mult decat cu oricare alte organisme si
capabili sa produca descendenti fertili, prin fecundare
reciproca".
George
Cuvier, in 1821, considera ca "trebuie
sa recunoastem neaparat ca anumite forme exista de la
inceputul lucrurilor, fara a depasi anumite limite si
ca toate fiintele apartinand uneia dintre aceste forme
constituie specia".
Jean Baptiste
Monet de Lamarck (1744 - 1829) avea, la inceput, convingerea ca "specia consta dintr-un numar de
indivizi asemanatori care, prin inmultire, dau indivizi
asemanatori lor" si ca "nu se poate contesta existenta speciilor in natura", ca
ulterior sa ajunga sa afirme: "Multa vreme am crezut ca in natura exista specii
constanteacum insa m-am convins ca aceasta a fost o
greseala si ca in natura nu exista decat indivizi".
Charles
Darwin (1809 - 1882) a ajuns la concluzia ca o "varietate bine pronuntata poate fi considerata ca fiind
o specie incipienta" si ca speciile nu sunt altceva decat "varietati bine pronuntate".
Emil
Racovita (1869 - 1947) defineste specia in numai patru cuvinte
(asa cum spune chiar el): orice "colonie
izolata de consangeni".
Si acum
multi biologi definesc specia pe baza a 1 - 2 criterii, de obicei
morfologice, definitorii in activitatea practica de delimitare a ei. Unul
dintre criteriile forte, la care apeleaza multi cercetatori este
"criteriul fiziologic" - adica incompatibilitatea sexuala a organismelor.
De pilda, in 1918, zoologul Stresemann aprecia ca formele care au
atins nivelul speciei diverg fiziologic intr-atat incat se pot intalni
fara a se incrucisa. Se impune, cu aceasta ocazie, o
precizare asupra careia vom reveni cand vom discuta problema implicatiilor
modalitatilor de reproducere asupra statutului speciei. Precizarea
este urmatoarea: daca absolutizam definitia
anterioara, atunci capata statut de specie orice forma
sezoniera, orice individ aberant (sub aspectul cromosomial de pilda),
orice hibrid, etc., izolat reproductiv de ceilalti indivizi ai speciei.
Tureson,
ecolog de renume, in 1929, da o definitie in conformitate cu care
specia este "o populatie ai
carei indivizi se pot incrucisa in natura dand o
descendenta viabila si fertila. Daca sunt
incrucisati cu indivizi apartinand altor populatii,
descendentii sunt cu viabilitate scazuta si in mare parte
sterili".
Th.
Dobzhansky, genetician si evolutionist, era de parere ca "specia reprezinta un stadiu al
procesului evolutiei in care grupurile de forme care alta data,
faptic sau potential, se incrucisau reciproc, au segregat in
doua sau mai multe manunchiuri de forme ce au devenit fiziologic
imposibil de a se incrucisa".
E. Mayer
sustinea ca "sunt specii acele
grupuri de populatii naturale care, faptic sau potential, se incruciseaza
intre ele si care, in inmultirea lor, sunt izolate de alte grupuri
analoage". Autorul si-a dezvoltat mai departe ideea precizand ca
specia noua ia nastere in cazul in care o populatie se
izoleaza de specia parentala si capata anumite
caractere ce contribuie la izolarea fiziologica sau o garanteaza.
La randul
nostru apreciem ca, in general, definitiile date speciei
accentueaza, in primul rand, caracterul de grup izolat, grup ce nu se
incruciseaza cu alte grupuri sau, daca totusi procesul se
produce, urmasii sunt de regula sterili. Prin urmare, in
conceptiile mentionate, acordandu-se mare importanta
factorului genetic - incompatibilitatea, cu determinism profund, intre diverse
informatii ereditare. Fara pretentia de exhaustivitate
propunem urmatorul mod de analiza a fenomenului speciei. Pe de o
parte specia reprezinta o etapa din organizarea si integrarea
materiei vii, etapa cu caracter obligatoriu si universal, in sensul
ca nici o entitate biologica de nivel individual nu poate exista in
afara speciei. In aceasta calitate, specia nu poate fi definita decat
prin trasaturile ei generale, unificatoare, valabile pentru orice
entiatate de acest rang, indiferent ca apartine microorganismelor,
plantelor sau animalelor, inferioare sau superioare. Prin urmare specia, ca
entitate biologica integratoare, cuprinde totalitatea sistemelor de ordin
individual, cu un anumit gard de similaritate, indiferent de gradul lor de
complexitate, specializare, integritate etc.
Pe de
alta parte, taxonomic judecand realitatea, in arborele filogenetic al
lumii vii exista o multitudine de specii, ca purtatoare si
rezultante ale procesului evolutiei, care delimiteaza sectiuni
clare si discrete, caracterizabile prin trasaturi delimitante
specifice. Din acest punct de vedere, o
specie nu poate fi definita decat ca totalitatea indivizilor care au
origine comuna, poarta o informatie ereditara omogena,
bine integrata si distincta (un genofond specific) si
ocupa un loc bine definit si particular in circuitul energetic
si material al ecosistemului si, implicit, al biosferei. Nefacand
nici o referire la modalitatea de reproducere, am extins categoria de specie
si la indivizii apomictici, excluzand totodata
dificultatile create de hibridarile interspecifice.
Nementionand explicit teritorialitatea, eliminam
dificultatile induse de speciile ubiquitare.
Din punct de
vedere sistematic, mai precis din punctul de vedere al conceptului de "specie biologica", prin prisma
caruia apartin aceleiasi specii doar indivizii interfertili,
unitatile taxonomice infraspecifice sunt (Zavadskii, 1968):
semispecia - o noua specie
aproape formata, o "specie tanara", o specie pe cale de a fi
complet izolata reproductiv de specia mama;
subspecia sau rasa geografica, inca nu are bariere reproductive cu
specia mama;
ecotipul, rasa ecologica
locala;
populatia
locala, subunitatea fundamentala din structura speciei (a carei
absenta, spre deosebire de subunitatile anterioare, este
imposibila);
ecoelementul
grupul morfo-biologic
biotipul
Ultimele trei
subunitati se constituie in componente ale populatiei.
Populatia ocupa locul central in infrastructura speciei, fiind
apta de existenta de sine statatoare pe timp nedefinit, are
un areal clar delimitat, are un rol si o pozitie stabile in
biocenoza, se caracterizeaza printr-un anumit nivel numeric, aflat in
echilibru dinamic etc.
Pe de
alta parte, desi interfertile, populatiile unei specii sunt in
mod obligatoriu alopatrice, cazuri de suprapunere a arealului fiind posibile
doar atunci cand intervine o izolare ecologica. Prin urmare, daca
formatiunile infrapopulationale sunt incapabile de a supravietui
de sine statator si independent, populatia reprezinta
prima structura intraspecifica apta de a se mentine, de a
se perpetua si de a evolua. Intr-o tentativa de definire a
populatiei se poate afirma ca aceasta reprezinta "forma de existenta a speciei,
posedand toate conditiile necesare existentei si
dezvoltarii de sine statatoare timp nelimitat si,
totodata, capabila de a reactiona adaptiv fata de
modificarile mediului extern" (Svart, 1969 - citat de Botnariuc,
1979).
Prin prisma
conceptiei sistemice, populatia reprezinta un sistem biotic
supraindividual, in esenta echivalent cu specia, care este
caracterizat prin toate trasaturile specifice unui sistem viu, fiind
definit, in plus si de mentinerea unui nivel numeric optim. Cu
aceasta ocazie se impune o precizare: nivelul numeric poate fi extrem de
variabil, de el depinzand si asigurarea heterogenitatii interne
a unei populatii si, de aici, gradul ei de integralitate. Echilibrul
numeric intr-o populatie reprezinta un parametru dinamic a carui
valoare optima se poate atinge in timp (mai indelungat sau mai scurt, in
functie de numarul de indivizi pe care se constituie populatia -
numarul initial de indivizi). De cele mai multe ori o populatie
poate lua nastere din unul sau mai multi indivizi care, din diferite
cauze, au emigrat sau au fost diseminati pasiv, dintr-o alta
populatie. Evident, pentru a se forma o noua populatie si
nu doar o simpla extindere a arealului celei vechi, este obligatoriu ca
indivizii "fondatori" sa nu mai aiba contact, sa nu mai
intretina flux genic cu populatia de origine. In acest fel
privita problema, se poate afirma ca, prin populatie "mendeliana" se intelege totalitatea
indivizilor unei anumite specii, intre care se poate realiza incrucisarea
sexuata, si care au ascendenti si descendenti comuni
(Raicu, 1980). Desigur, nu putem sa nu remarcam ca, prin
aceasta definitie sunt excluse din categoria populationala
sistemele a caror perpetuare si mentinere a nivelului numeric sunt
asigurate prin reproducere apomictica. Intrucit, sistemic abordand
problema, etapa de organizare si integrare populationla
vizeaza intreaga materie vie de pe Terra (nu exista individ biologic
care sa nu fie sau sa nu devina integrat intr-o populatie),
apreciem ca o definitie a populatiei trebuie sa fie
universal valabila si nu exclusivista. In consecinta,
consideram ca o
populatie, atat ca unitate sistematica, cat si ca etapa din
organizarea si integrare a lumii vii, reprezinta totalitatea
indivizilor care au origine comuna, contribuie la asigurarea unui fond
integrat de gene pe un areal discret si comun, constituindu-se intr-un
ansamblu unitar, cu rol bine definit in fluxul material si energetic din
cadrul sistemului imediat urmator (biocenoza) de integrare. In acest
context dispare obligativitatea reproducerii sexuate ca o conditie de
baza a existentei unei populatii. Si in cazul
populatiilor apomictice informatia ereditara circula intre
indivizii generatiilor succesive (intre generatiile parentale si
cele filiale). Prin urmare nu se poate afirma ca se produce o intrerupere
a fluxului informational interindividual. Mai mult, atat intr-o
populatie amfimictica (panmictica mare), cat si in una
apomictica, exista un fond de gene a carui evolutie, in
sensul schimbarii raporturilor procentuale intre gene si alele, nu
depinde doar de modalitatea de reproducere ci, in primul rand de presiunea
selectiei.
Cook (1976)
precizeaza ca populatia reprezinta un grup de organisme in
spatiu, reprezinta totalitatea
indivizilor interancrucisabili. Zavadskii (1968), la randu-i,
considera populatia locala drept unitatea fundamentala de
existenta, adaptare si reproducere a speciei. Nu exista o
interdependenta clara intre dimensiunile arealului si
numarul populatiilor locale. Uneori specia este continua, iar
limitele populatiei sunt strans legate de calitatea sa de unitate
reproductiva. In acest sens trebuie sa admitem faptul ca, la
speciile cu reproducere sexuata, cea mai mica unitate reproductiva
o constituie perechea de indivizi unisexuati, sau chiar un individ
hermafrodit. Populatia locala, ca unitate reproductibila,
cuprinde o multitudine de indivizi inruditi, dar deosebiti genotipic,
intre care intervin, constant, incrucisari libere si nelimitate
(panmixie). Schimbul nelimitat de gameti favorizeaza
inmagazinarea in fiecare individ a
potentelor genetice ale intregii populatii. De aceea, separarea unor
indivizi (sau perechi) din cadrul populatiei si inmultirea lor
aparte, duce la reducerea considerabila a capacitatilor
biologice ale descendentilor si la pierderea plasticitatii
ecologice. Populatia locala
este o comunitate cimentata intr-un intreg prin procesele de
incrucisare. De aceea, pentru a caracteriza specia ca unitate
autoreproducatoare, sunt extrem de importante datele privind limitele
precise ale populatiei locale, ca si cele privind hotarele dincolo de
care inceteaza orice schimb de gameti (intre populatii
diferite). Specia este deci reprodusa
de populatiile locale. Populatia locala, pe de alta parte,
reprezinta o formatiune care poseda capacitatea reala de a
atinge limita inferioara a nivelului de specie (nivel de organizare
si integrare a lumii vii).
Cand se
determina limitele populatiilor locale trebuie sa se
tina seama de gradul de sedentarism al indivizilor sau familiilor de
indivizi. In context, trebuie elucidata notiunea de raza de actiune, notiune
care implica alte notiuni ecologice particulare ca: zona de vanat, zona de crestere a puilor, zona de inmultire etc. La
plante, raza de actiune a individului poate fi egala cu distanta
la care se extinde sistemul radicular si partea aeriana, cat si
cu distanta la care actioneaza substantele biologic active
eliberate de individ. Raza de actiune a individului creste in cazul
in care la reproducere participa sporii, polenul, semintele etc. La
animale raza de actiune este influentata de sedentarism, dar
si de faptul ca idivizii se intorc mereu in acelasi loc, chiar
de la distante foarte mari (anguilele, unele pasari, unii fluturi
etc.).
In
biosfera se intalnesc toate gradele de trecere de la specii
unipopulationale la specii divizate in numeroase populatii, perfect
izolate. Evident, speciile care au perspectivele cele mai mari de evolutie
sunt cele care detin populatii vaste, subampartite, la
randul lor, in multe populatii mici (micropopulatii) si numai
partial izolate reproductiv. Datorita izolarii incomplete, toate
proprietatile utile care apar in cadrul unei micropopulatii, pot
deveni repede un bun al intregii populatii. Datorita unui anumit grad
de izolare, unele modificari adaptative vor avea tendinta de a se
acumula in interiorul unei micropopulatii izolate. Existenta
schimbului de gameti ingaduie raspandirea adaptarilor de
insemnatate generala in cadrul intregii populatii.
Ingradirile vizand acest schimb ceeaza posibilitatea acumularii
adaptarilor de semnificatie particulara numai in limitele unei
micropopulatii.
In cazul
inmultirii apomictice unitatea reproducatoare este individul.
Descendentii unui singur individ, autogam sau apomictic (clona),
reprezinta o multitudine de indivizi inruditi care au un genotip
omolog dar nu reprezinta o unitate autoreproducatoare. Deosebirea
dintre clona sau populatie clonala alcatuita din
cateva linii clonale si populatia locala panmictica,
consta in faptul ca o clona reprezinta o suma de indivizi
ce se autoreproduc, iar populatia amfimictica (ca intreg)
reprezinta o unitate autoreproductibila. Tocmai aceasta
deosebire principala intre intre unitati elementare
autoreproducatoare constituie cauza dificultatilor particulare
care intervin in procesul
delimitarii hotarelor speciilor, la formele agame ale organismelor
inferioare, ca si la plantele superioare care adopta reproducere
apomictica. Desigur ca, la grupurile cu inmultire asexuata,
capacitatea de autoreproducere nu reprezinta un caracter atat de dezvoltat
al populatiei considerata unitate integrala, incat sa
poata fi inclus in randul caracterelor principale. La autogame structura
populatiei poate fi considerata intermediara intre
populatiile amfimictice tipice si populatiile clonale tipice.
Daca autogamia este reala, populatia reprezinta un ansamblu
de linii coexistente. Dar, in aceste populatii actioneaza
si un factor integrator (intr-o oarecare masura), datorat
fecundarii incrucisate. In fapt, gradul de integrare al acestor
populatii de plante poate fi influentat, intrucatva, de
frecventa incrucisarilor interindividuale. Dar, un rol la fel de
important il au si interactiunile dintre indivizi, prin intermediul
substantelor bioactive, asa ca populatiile clonale nu sunt
doar amestecuri mecanice de forme mai mult sau mai putin
asemanatoare.
In
functie de dimensiuni, adica in functie de nivelul numeric,
populatiile pot fi clasificate in: populatii
mici (cu mai putin de 100 de indivizi), populatii mijlocii (care cuprind cateva sute de indivizi), populatii mari (cu cateva mii de
indivizi) si populatii foarte
mari (cu peste 10.000 de indivizi cu capacitate de reproducere).
Fara indoiala ca, din punct de vedere genetic, marimea
populatiei are o foarte mare importanta deoarece isi pune
amprenta asupra legilor geneticii populationale, legi cu caracter
probabilistic. Or, o lege cu caracter probabilistic se realizeaza cu atat
mai exact cu cat o populatie are un efectiv mai mare.
Mai trebuie
precizat ca desi izolarea interpopulationala
reprezinta o conditie sine qua
non a delimitarii si existentei unei populatii,
schimbul de informatii ereditare (fluxul de gene) intre populatii,
mai apropiate sau mai indepartate, nu este exclus. Schimbul (in sensul
fluxului) de gene se poate realiza fie pri intermediul gametilor si
al indivizilor, fie numai prin intermediul indivizilor, in functie de
modalitatea de reproducere (amfimictica, in primul caz, apomictica,
in cel de-al doilea caz). Lipsa panmixiei caracterizeaza atat
populatiile plantelor cultivate cat si pe cele ale animalelor
domestice, indiferent daca sunt amfimictice sau apomictice, deoarece omul
controleaza si dirijeaza incrucisarile
interindividuale.
O
pozitie intermediara intre populatiile apomictice si cele
amfimictice o ocupa populatiile alcatuite din indivizi autogami.
Fenomenul este, desigur, mai mult frecvent la plante. De pilda, intr-o
populatie autogama un individ de tipul AA (homozigot dominant)
formeaza numai gameti de tipul A si descendenti in
exclusivitate de tipul AA. In cazul unor indivizi monozigoti pentru mai
multe perechi de gene, cum ar fi de pilda AabbccDD, ,gametii vor fi
de tipul AbcD. Descendenta va fi, deasemenea, identica si
monozigota. Astfel stand lucrurile este evident ca, din punct de
vedere al comportarii fondului de gene, intre autogamie si apomixie
nu exista diferente. Diferente apar doar in cazul in care,
intr-o populatie, se gasesc mai multe tipuri de homozigoti care,
intamplator, uneori, pot sa se incruciseze. De exemplu, daca
exista indivizii AABB si aabb, printr-o eventuala hibridare
rezulta AaBb. In cazul autofecundarii, alaturi de tipurile
parentale apar si noi combinatii de gene si, deci de caractere
de tipul aaBB si Aabb. Fara indoiala procesele au cu totul
alta dinamica in cazul alogamiei si este cunoscut faptul ca
speciile animale sunt prin excelenta alogame si, in plus, au
si o mare mobilitate.
O
populatie alogama (panmictica deci) este constituita din
indivizi heterozigoti, caracterizati printr-o mare variabilitate,
atat genotipica, cat si fenotipica. In plus, prin incrucisarea
continua intre organisme heterozigote pentru un numar mare de gene au
loc noi recombinari, in fiecare generatie, si amplitudinea de
variabilitate poate atinge valori din ce in ce mai mari. Tinand cont de
faptul ca, uneori, genotipul unui individ poate contine multe mii de
gene cu alelele lor, precum si de posibilitatea ca heterozigotia
sa se manifeste pentru 1,2,3,n loci, se poate aprecia ca intr-o
populatie oricat de mare sunt conditii ca sa nu existe doi
indivizi la fel. Reamintim doar fenomenul urmator: numai prin
distributia intamplatoare a cromosomilor omologi in gameti, prin
intermediul meiozei, exista sansa formarii a 2n
gameti diferiti, de catre fiecare individ (n reprezentand ½ din numarul de factori ereditari din celula
somatica). Prin intalnirea a 2n gameti masculi cu 2n
gameti femeli, pot rezulta 2n gameti diferiti intre
ei, doar de la un singur cuplu parental. Dar, intr-o populatie exista
nenumarate cupluri, interschimbabile de la o generatie la alta
si, in consecinta, variabilitatea este nelimitata.
Daca o populatie panmictica ar perpetua prin consagvinizare, in
timp, s-ar ajunge la "desfacerea" respectivei populatii in nenumarate
linii pure. De pilda, in cazul autogamiei, admitand ca avem un
individ heterozigot intr-un singur locus (Aa), el va forma doua tipuri de
gameti A si a, in proportie egala. Prin intalnirea
probabilistica a acestor gameti (probabilistica este aceeasi
pentru intalnirile AA, Aa si aa), rezulta trei tipuri de indivizi,
reprezentand tot atatea genotipuri.
Prin urmare,
genetica populatiilor presupune studiul comportamentului genelor in
populatii, in contextul adaptarii organismelor la medii de
viata stabile sau schimbatoare. Altfel spus, genetica
populatiilor se implica in studiul mecanismelor evolutiei,
acceptat fiind faptul ca evolutia
reprezinta, printre altele, un proces de continue schimbari in
frecventa genica. In consecinta, preocuparea centrala a geneticii populatiilor consta in
investigarea dinamicii modificarii frecventei genice. Dealtfel
este bine sa precizam ca genetica populatiilor este
diferita de orice alta parte a geneticii. De aceea este indicat
sa se recurga, inca de la inceput, la elucidarea si
edificarea terminologiei specifice.
III.2. Factorii
care determina compozitia si dinamica genetica a populatiilor
Echilibrul
genetic in populatii, demonstrat pe baza analizei genetice a structurii
populatiei, este posibil sa se mentina constant numai in
cazul absentei actiunii modificatoare a unor factori. Se stie
ca in natura, asupra populatiilor actioneaza permanent
numerosi factori, care pot duce la schimbarea echilibrului genetic
anterior, fie prin pierderea unor genotipuri, fie prin aparitia unor
genotipuri noi in populatie.
Aceasta
inseamna ca de la o generatie la alta fondul genetic al unei
populatii se gaseste intr-un echilibru dinamic determinat de
factori, ca: mutatia, migratia, selectia, driftul genetic,
conditiile de existenta, etc.
Efectele mutatiei in populatii. Mutatiile
reprezinta o sursa importanta de variabilitate genetica a
organismelor, contribuind ca un factor pozitiv la evolutia acestora.
Efectul mutatiei asupra fondului genetic al unei populatii este
sensibil in cazul in care procesul mutagen are un caracter permanent. Cand
procesul mutagen are loc sporadic, efectul mutational in populatie
devine insensibil. De aceea, mutatiile care apar in populatii pot fi nerecurente si recurente.
Mutatiile nerecurente se caracterizeaza prin
aceea ca nu au capacitatea de a produce o schimbare permanenta in
fondul genetic al populatiei. Sa presupunem ca procesul mutagen
afecteaza numai un singur reprezentant dintr-o intreaga
populatie. O asemenea mutatie este de mica importanta
pentru a provoca o schimbare sensibila a frecventei genei,
datorita faptului ca o singura mutatie dispune de o
sansa minima de a se mentine intr-o populatie mare,
mai ales cand aceasta nu constituie un element pozitiv in procesul
selectiei.
In
conformitate cu legitatea frecventei genelor in populatie,
probabilitatea ca o gena sa se mentina viabila
descreste de la o generatie la alta, pana ce se reduce la
frecventa zero in populatia respectiva. Asemenea mutatii,
lipsite de avantajul selectiv si deci, fara viabilitate in
populatie, sunt cunoscute sub forma de mutatii nerecurente.
Mutatiile recurente, spre deosebire de cele
nerecurente, provoaca schimbari in frecventa genelor. Aceasta
inseamna ca mutatia recurenta este rezultatul unui proces
mutational care are loc cu regularitate, asigurand o frecventa
ritmica a genelor, astfel ca fiecare gena mutanta nu va mai
fi predispusa la disparitie.
Daca in
populatiile A1A2, gena A1 sufera un proces de mutatie in A2, in
populatia respectiva se va reduce numarul de gene A1
si vor aparea gene A2. De asemenea, prin mutatie de
reversie, genele A2 pot sa muteze in A1.
Fata de aceste posibilitati de mutatie, si anume
A1 in A2 si A2 in A1, devine
necesara stabilirea raportului frecventelor celor doua gene A1
si A2, in cadrul populatiei respective, fiind un raport
exprimat printr-un echilibru.
In vederea
determinarii acestui raport, se folosesc urmatoarele elemente de
calcul: q - frecventa genei in populatie; u - proportia
mutantelor A1 in A2; v - proportia mutantelor A2
in A1.
Daca
admitem ca proportia mutantelor A1 si A2
este de 2/1000 gameti, adica u = 0,002; iar proportia mutantelor
A2 in A1 de 1/1000, adica v = 0,001, se poate
stabili raportul de participare in populatie a celor doua gene (A1si
A2).
Intrucat
raportul care se creeaza intre frecventele genelor ce alcatuiesc
genofondul populatiei se concretizeaza intr-un echilibru,
inseamna ca numarul de gene care muteaza intr-un sens va fi
egal cu cel care muteaza in sens invers.
In acest caz,
pentru a afla frecventa genei A1, deci proportia
mutantelor A2 in A1, se foloseste formula:
q
= v/(u+v).
Pentru q (A1) = 0,001/(0,002+0,001) = 0,001/0,003 = 0,33
Exprimata
in procente, frecventa mutantelor A2 in A1(deci a
genei A1) este egala cu 33%. Pentru realizarea unui echilibru
intre genele A1 si A2, rezulta ca gena A2
va avea frecventa egala cu 67%, adica cu diferenta (100 -
33 = 67).
Exista
insa si cazuri cand frecventa mutatiei genei intr-un sens
nu se sincronizeaza cu mutatia celeilalte gene in sens invers,
datorita unei lupte pentru existenta care apare intre
componentii populatiei. O asemenea lupta pentru existenta
intre componentii unei populatii, exprimati prin genotipuri
diferite, a fost semnalata de catre Ph. L Heritier si G. Teissier. Ei au introdus intr-o
populatie mutanta de Drosophila
melanogaster cu ochiul "bar" cateva perechi de Drosophila normala (salbatica). Dupa 100 de
zile au constatat ca numarul insectelor cu ochiul "bar" a scazut
cu 40%. Dupa 500 de zile au observat disparitia totala a
indivizilor cu ochiul "bar".
Rezultatul
ca in cazul unei lupte pentru existenta intre cele doua
genotipuri, echilibrul nu se stabileste prin mutatia de reversie, ci
prin dominanta totala a drosofilei normale care s-a impus in procesul
selectiei.
Efectele migratiei in populatie. Prin migratie se
intelege miscare genelor si genotipurilor intre populatii
diferite. In populatiile naturale de plante, migratiile implica
transferul de polen sau de seminte. Transferul de polen corespunde unui
transfer de gameti, iar cel de seminte corespunde unui transfer de
zigoti.
Migratia
cuprinde doua laturi, si anume: emigrarea
unor gene, datorita transferului de polen sau de seminte in afara
populatiei, intr-o alta populatie, si imigrarea de gene, adica aducerea de gene noi, straine,
in componenta unei populatii.
Infiltrarea
unor gene noi prin intermediul polenului, si deci a gametilor, ca
urmare a participarii gametilor straini la fecundarea
gametilor dotati cu gene proprii populatiei respective, duce in
mod implicit la transmiterea genelor noi generatiilor urmatoare
si deci, la o imbogatire a fondului genetic al populatiei.
Pentru a
evalua cantitativ efectul migratiei in populatie, consideram
ca proportia genelor provenite prin migratie (imigrare) este
egala cu m, iar restul de 1 - m o reprezinta genele native ale
populatiei respective. Daca notam frecventa unei gene
oarecare cu qm pentru genele imigrante si cu qo
pentru genele native, frecventa unei gene raportate la intreaga
populatie notata cu q1 va fi data de formula:
q1
= mqm + (1 - m)qo = m(qm - qo ) + qo
Intrucat
frecventa unei gene imigrante se modifica dupa fiecare
generatie de imigrare, inseamna ca aceasta imigrare poate fi
stabilita pe baza diferentei dintre frecventa dinainte de
imigrare si frecventa de dupa imigrare, diferenta care
se noteaza cu Δq. In acest caz, Δq = q1 - qo =
m (qm - qo).
Din
aceasta ecuatie deducem ca ritmul in care are loc schimbarea
frecventei unei gene, intr-o populatie supusa imigrarii
depinde de ritmul de imigrare si de diferenta dintre frecventa
genelor imigrante si a celor native. Efectul migratiei se manifesta in raport invers cu efectivul
populatiei, in sensul ca este mai sensibil in populatiile mici.
Din punct de
vedere practic, efectul migratiei poate fi valorificat atunci cand genele
noi ajunse in populatie, duc la o imbunatatire a structurii
genetice a populatiei. In cazul in care se urmareste
pastrarea in stare pura a materialului biologic (a fondului genetic)
se iau masuri de evitare a imigrarii unor gene, prin izolarea
plantelor, astfel ca graunciorii de polen straini sa nu fie
adusi de vant sau insecte de la o populatie la alta.
Efectele selectiei in populatii. Selectia
reprezinta procesul de supravietuire a organismelor, ale caror
genotipuri le asigura cea mai mare capacitate de adaptabilitate
fata de conditiile de mediu in cursul vietii individuale
si, ca urmare, perpetuarea prin cei mai vigurosi descendenti.
Posibilitatea
organismelor de a supravitui si de a da urmasi este
conditionata de gradul de adaptare al descendentilor la
aspectele variate ale mediului, exprimandu-si prin aceasta capacitatea lor
selectiva, care reprezinta factorul primordial al dinamicii
populatiei. De asemenea, aparitia de noi genotipuri in cadrul unei
populatii reprezinta rezultatul actiunii selectiei
naturale. Exista si cazuri cand actiunea selectiei asupra
genotipurilor poate duce la pastrarea acestora, la diminuarea lor
numerica sau chiar la disparitia lor.
Se poate
aprecia masura in care se manifesta actiunea selectiei cu
ajutorul a doi indicatori: valoarea selectiva si coeficientul de
selectie.
Valoarea selectiva. Este sinonima cu valoarea adaptativa a genotipurilor. Se
stie ca selectia actioneaza diferit asupra
mutatiilor, in functie de starea lor dominanta sau
recesiva. Astfel, cu cat un genotip realizeaza o mai buna
capacitate de supravietuire, in sensul unei fecunditati
ridicate, cu atat intr-o populatie el este exprimat printr-o mai mare
raspandire. Asemenea genotipuri dispun de o valoare selectiva
ridicata.
De aceea, valoarea selectiva a unui genotip
poate fi evaluata prin numarul de descendenti pe care-l
realizeaza, fata de numarul de urmasi produsi de
genotipurile normale, considerand ca valoarea selectiva a acestora
este de 100%, adica egala cu unitatea.
Coeficientul de selectie (S). Analiza genetica a unei
populatii da posibilitatea sa se determine valoarea
selectiva a diferitelor genotipuri. Daca in urma unei analize
genetice se constata ca indivizii homozigoti cu genotipul aa
sunt in numar mai mic decat cei reprezentati de formele dominante cu
genotipurile AA si Aa, si anume 80 de indivizi cu aa, fata
de 100 cu AA sau Aa, se poate afla valoarea coeficientului de selectie a
homozigotilor recesivi.
In acest caz,
se considera ca valoarea selectiva, ca termen de
comparatie, o constituie valoarea selectiva a dominantelor (AA
si Aa), care este egala cu 1, respectiv 100%. Valoarea selectiva
a homozigotilor recesivi fiind de 0,8 (80%) inseamna ca
diferenta dintre aceste doua valori exprima tocmai coeficientul
de selectie (S)
S
= 1 - 0,8 = 0,2
Valoarea
coeficientului de selectie (S) indica procentul descendentilor
dintr-un anumit genotip care se elimina din populatie ca urmare a
actiunii selectiei. In cazul de fata, indivizii cu
genotipul aa se elimina intr-o proportie de 20%. Rezulta ca
valorile S pot fi cuprinse intre 0 (zero) si 1.
Atunci cand
genotipurile care se compara au aceeasi fecunditate si procent
de supravietuire, deci cu valoare selectiva normala, valoarea
coeficientului de selectie va fi:
S
= 1 - 1 = 0
In acest caz,
toti indivizii din populatie au sanse de supravietuire
egale.
Daca in
populatie un anumit genotip homozigot recesiv provoaca letalitate sau
sterilitate, inseamna ca valoarea sa selectiva va fi egala
cu 0 (zero), iar eliminarea lui integrala din populatie este
iminenta. Exprimarea matematica a acestui caz rezulta din
ecuatia: S = 1 - 1 = 0.
Genofondul
unei populatii, datorita actiunii selectiei, poate fi
complet purificat prin eliminarea genelor nefavorabile, dominante si
recesive. Eliminarea este conditionata de marimea
populatiei, si anume in
populatiile mici se mareste posibilitatea unirii gametilor
cu gene identice, si deci, se mareste frecventa formelor
homozigote. In acest fel, selectia elimina mai repede genele
daunatoare si cumuleaza pe cele folositoare.
In ceea ce
priveste ritmul eliminarii genelor nefavorabile, ca urmare a
actiunii selectiei se constata ca organismele care
poseda gene letale dominante sau o sterilitate completa sunt
eliminate inca din prima generatie, in timp ce genele dominante cu o
expresivitate mai mica se elimina treptat din populatie.
Datorita mutatiilor care apar iau nastere gene
asemanatoare celor eliminate si astfel devine imposibila
disparitia acestora.
Driftul genetic in populatiile mici. In populatiile
prevazute cu un numar redus de indivizi, modificarea frecventei
genelor este posibila fara interventia selectiei, ca
urmare a unor imprejurari intamplatoare care afecteaza procesul
reproducerii.
Fenomenul de
crestere sau de scadere brusca a frecventei genelor in
populatii mici, datorat unor factori neprevazuti, poarta
numele de drift genetic.
Pentru a
intelege actiunea driftului genetic in populatiile mici folosim
de exemplu incrucisarea intre doua soiuri de grau nearistate,
heterozigote, cu formula genetica Aa. Factorul A determina caracterul
mutic (lipsa aristelor), iar factorul a determina prezenta aristelor.
Fiecare din cei doi genitori formeaza gameti (masculi si femeli)
atat cu factorul A cat si cu factoru a. In cazul in care in urma
incrucisarii numarul descendentilor este mare, segregarea
are loc in raportul de 25% homozigoti cu genotipul AA, 50%
heterozigoti Aa si 25% homozigoti aa. La un numar redus de
descendenti, deci dintr-o populatie mica, imprejurari
neprevazute, cauzate de factori de mediu, pot provoca fecundarea numai a
gametilor cu factorul a, prin distrugerea celor cu factorul A. In acest
caz se obtin numai plante aristate (aa), ca urmare a pierderii factorului
A continut, de pilda, in anumiti graunciori de polen.
Intrucat
driftul genetic este prezent numai in populatiile mici, este necesar a
lucra cu efective cat mai mari, in scopul evitarii efectelor negative ce
survin de pe urma numarului restrans de indivizi, folosit ca material de
selectie. In populatiile mari, frecventa unei gene este
insesizabila, deoarece cresterea sau descresterea numerica
a acesteia se echilibreaza in interiorul populatiei respective prin
alela sa.
Conditiile de existenta ca factor al
modificarii structurii populatiei.
Structura
genetica a populatiilor poate sa sufere modificari, atat
datorita unor conditii ale mediului abiotic, cum sunt: hrana,
umiditatea, temperatura, lumina etc. Cat si datorita unor
conditii biotice, determinate de relatiile dintre organisme.
In
functie de gradul homozigotiei sau heterozigotiei indivizilor
care alcatuiesc populatia, s-a constatat ca populatiile
autogame sunt mult mai stabile, in comparatie cu cele heterozigote, care
manifesta o mare labilitate.
Stabilitatea
populatiilor autogame se traduce printr-o capacitate mai redusa de
adaptare la conditiile de mediu schimbate. Populatiile allogame
manifesta o mare capacitate de adaptare la conditiile variate ale
mediului, deoarece heterozigotia le confera insusirea de
labilitate.
In general,
factorii climatici afecteaza structura genetica in dinamica
evolutiei populatiilor, ca urmare a duratei prelungite a
fluctuatiilor conditiilor de clima, a carei
influenta se concretizeaza in schimbari ale mecanismelor
fiziologice si de comportament si deci, si a mecanismului
genetic.
Aparitia
unor bariere naturale, de netrecut, pe arealul unei populatii, cauzate de
modificari ale reliefului, de unele fenomene geologice, prezenta
fluviilor, mlastinilor etc., limiteaza incrucisarile libere
ale indivizilor in cadrul unei specii sau populatii.
Climatul
teritorial, climatul sezonier, microclimatul, ca elemente ale unei izolari
ecologice, favorizeaza dezvoltarea unor anumite genotipuri in
componenta populatiei.
Incrucisarile
libere in cadrul populatiei mai pot fi limitate si de factori
genetici, exprimati prin fenomenul de poliploidie, prin aparitia
diferitelor aberatii cromosomale, prin existenta unor
incompatibilitati nucleo-citoplasmatice, marirea numarului
de gene letale si a mutatiilor
sterile etc., care la un loc reprezinta cauze ale unei izolari
genetice si fiziologice.
Homeostazia genetica in populatii. Prin homeostazie genetica se intelege
efectul tuturor proceselor adaptative care asigura unei populatii
capacitatea de a-si pastra structura genetica fata de
actiunea factorilor din mediul extern. Adaptarea unei populatii se
realizeaza pe seama heterozigotiei, polimorfismului, driftului
genetic, influentei conditiilor de existenta,
polenizarii selective in cadrul populatiei si a altor factori.
Heterozigotia in populatii reprezinta
unul din mecanismele principale ale homeostaziei genetice fiind subliniata
de I. M. Lerner (1954), Emerson (1952), J. B. S. Haldane (1955), pe baza
efectelor sale de supradominanta. In general, orice populatie
panmictica ascunde o diversitate genetica, determinata de de un
procent ridicat de heterozigotie si de diferite mutatii.
Pornind de la
legea unitatii biologice a
incrucisarii, enuntata de Ch. Darwin, s-a ajuns la
concluzia ca starea de heterozigotie a genelor allele duce la o marire
a vigorii organismelor hibride si a dezvoltarii
posibilitatilor de adaptare. Incrucisarile inrudite si
autofecundarea repetata a plantelor allogame, intensifica
homozigotia la descendenti, in sensul ca, acestia
manifesta fenomenul de degenerare care le diminueaza capacitatea de
adaptare la conditiile de mediu.
Formele
mutante, recurente, in cadrul populatiei, sporesc starea de
heterozigotie si contribuie, de asemenea, la marirea
capacitatii de adaptare a populatiei.
Polimorfismul ereditar, bazat pe existenta in
aceeasi populatie a unor diversitati genotipice,
reprezinta un alt mecanism genetic adaptiv, care asigura unitatea
genetica a unei populatii. Prezenta polimorfismului si a
diferitelor moduri de manifestare se datoresc actiunii selectiei
naturale.
Efectele
tuturor mecanismelor de adaptare a populatiilor se concretizeaza in
diversificarea structurii lor genetice, ca urmare a actiunii selective a
conditiilor de existenta in diferentierea genetica a
populatiilor.
Mentinerea
sau schimbarea echilibrului genotipurilor in populatii reprezinta un
proces continuu de adaptare a acestora la conditiile variate ale mediu.
Fenomenele de mutatie si selectie, care ocupa un loc
important in studiile de genetica a populatiior, contribuie la
explicarea capacitatii de evolutie a organismelor constituite in
populatii.
Faptul
ca pe arealul de raspandire al populatiilor pot surveni anumite
obstacole, ca factori de izolare a populatiilor se creeaza
posibilitatea de evolutie divergenta, care poate sa aiba
drept consecinta, diferentierea populatiilor in subspecii
sau chiar specii.
IV. SCOPUL INVESTIGATIILOR
Exista parerea, falsa, ca Drosophila melanogaster, ca material
pentru investigatiile de ordin genetic, a cazut in desuetudine. Ne
permitem sa afirmam ca lucrurile nu stau deloc asa,
deoarece, consultand literatura de specialitate, de ultima ora,
gasim o multitudine de cercetari, cu tematica de
citogenetica, genetica populatiilor, mutageneza si chiar
genetica moleculara, care au ca obiect de studiu faimoasa, de acum,
musculita a otetului. In consecinta nu am ezitat cand,
pusi in fata alegerii materialului biologic pentru
investigatiile ce ni le-am propus, am optat pentru Drosophila melanogaster.
Desi
elucidate in principiu, mai ales la procariote, mecanismele reglajului genetic
sunt inca foarte putin aprofundate la eucariote. Un alt aspect
insuficient investigat vizeaza stabilirea cu exactitate a
corelatiilor complexe dintre fenotipizarea concreta a
informatiei genelor si alte procese ce au loc pe parcursul
ontogenezei. Si investigatiile de acest tip au ca obiect Drosophila melanogaster, desigur,
alaturi de alte specii.
In lucrarea de fata ne-am propus, ca etapa
in caracterizarea complexa a unor populatii naturale si
experimentale de Drosophila melanogaster,
studiul dinamicii natalitatii si a ratei sexelor, studiul
actiunii selectiei naturale pe parcursul a doua sau trei
generatii succesive, in populatii naturale si
micropopulatii experimentale, sub efectele consangvinizarii si
tratamentelor cu factori mutageni fizici si chimici.
In cadrul experimentului in cre s-au urmarit efectele
consangvinizarii in doua populatii locale de Drosophila melanogaster, au fost
propuse studiul dinamicii
natalitatii, ratei sexelor si a duratei eclozarii pontei,
in trei generatii succesive, in doua populatii naturale.
In ceea ce priveste efectul tratamentului cu nicotina, tratatamentele cu factori xenobiotici, administrate
indivizilor unor specii animale si/sau vegetale, furnizeaza date
deosebit de interesante si, uneori, utile, relative la efectele
stimulatoare, inhibitoare sau mutagene. Pentru activitatile cu
caracter practic, mai ales cele de selectie si ameliorare,
detinerea unui material initial cu o larga amplitudine de variabilitate,
reprezinta o conditie "sine qua non". De aceea consideram
ca acest tip de investigatii isi pastreaza, inca,
toata importanta si valoarea. In lucrarea de fata
ne-am propus investigarea actiunii nicotinei, in diferite concentratii,
asupra unei populatii de Drosophila
melanogaster in care a avut loc
reversmutatia pentru locusul eyeless. Datorita faptului ca in
populatia respectiva apareau indivizi, intr-un procent destul de
constant, ce prezentau diferite abateri de la forma normala a ochilor
si a aripilor, am considerat importanta investigarea modificarii
acestui raport in urma tratamentului cu nicotina.
V.
MATERIALE SI METODE DE LUCRU
V. 1. Efecte ale consangvinizarii in doua populatii locale de Drosophila melanogaster
Materialul si metoda de lucru
Investigatiile au vizat
indivizi din populatiile Botosani si Suceava. S-au luat cate 10
cupluri (♂ + ♀), repartizate in 10 fiole cu mediu de cultura,
carora li s-a analizat descendenta, pe trei generatii, sub
aspectul prolificitatii, ratei sexelor, duratei eclozarii pontei
si a aparitiei eventualelor mutatii. De la o generatie la
alta, perpetuarea s-a asigurat prin incrucisari de tip frate x sora.
V.2. Efecte ale tratamentului cu nicotina, in diferite
concentratii, aplicat indivizilor de Drosophila
melanogaster dintr-o populatie reversmutanta
Materialul si metoda de lucru
Pentru a
testa efectele diferitelor concentratii de nicotina, administrate
indivizilor de Drosophila melanogaster s-a procedat astfel:
Nicotina a
fost administrata in concentratiile de 0,005%, 0,01%, 0,03%. Pentru
fiecare concentratie experimentul s-a repetat de trei ori. In cadrul
fiecarei repetitii s-au luat cite trei eprubete, in fiecare
eprubeta s-au introdus doi masculi si doua femele de Drosophila
melanogaster apartinand unei linii
reversmutante din Laboratorul de Genetica. Mutatia a carei
reversie s-a produs, era eyeless (indivizi fara ochi). In literatura
de specialitate este mentionat faptul ca respectiva mutatie
cunoaste un grad inalt de reversie (este foarte instabila).
V.3. Dinamica fondului de gene in populatii experimentale de Drosophila melanogaster sub presiune
selectiva
Materialul
si metoda de lucru
Pentru a
evidentia dinamica fondului de gene intr-o populatie
experimentala de Drosophila
melanogaster , sub presiune selectiva, s-a procedat astfel:
Intr-un
flacon cu mediu de cultura s-au introdus cate cinci masculi si cinci
femele din urmatoarele linii din Laboratorul de Genetica: Oregon
(70), Brown (5), Ebony (46), Vestigial (46) si White (111).
Modificarile
vor aparea ca urmare a factorului de stres - suprapopulare.
Se va
urmari indeaproape evolutia fiecarei linii din cele participante
la experiment.
Experimentul
se desfasoara pe parcursul a doua generatii.
Experimentul
va fi repetat respectandu-se preocedurile folosite anterior, fiind
inlocuita linia mutanta Brown (5) cu Curly (10).
VI. REZULTATE SI DISCUTII
VI.1. Efecte ale consangvinizarii in doua populatii locale de Drosophila melanogaster
Experimentul a fost efectuat
in Laboratorul de Genetica a Universitatii "Alexandru Ioan Cuza"
din Iasi, in perioada 18 februarie - 2 aprilie 2002, prima generatie
fiind obtinuta intre 25.03.2002 si 02.04.2002, cea de-a doua
intre 05.03.2002 si 14.03.2002, iar cea de-a treia intre 25.03.2002
si 02.04.2002, pentru ambele populatii luate in studiu. In ceea ce
priveste populatia Botosani, pentru prima generatie, durata
de aparitie a tuturor indivizilor a fost de 6,9 zile, pentru a doua
generatie de 10 zile si pentru a treia generatie de 9 zile. Prin
urmare putem aprecia ca, sub efectul consangvinizarii s-a constatat o
usoara lungire a perioadei necesare eclozarii oualelor
si aparitiei tuturor descendentilor. La populatia Suceava,
durata a fost de 6,4 zile pentru prima generatie, de 9,6 zile pentru cea
de-a doua generatie si de 6,8 zile pentru cea de-a treia
generatie, de aceasta data concluzia de la prima populatie
fiind aplicabila doar pentru generatia a doua.
In ceea ce priveste
prolificitatea per cuplu in populatia Botosani, in prima
generatie a fost de 81,3 indivizi, in cea de-a doua generatie a fost
de 136,6 indivizi, iar in cea de-a treia generatie a fost de 68,5
indivizi. S-a putut calcula ca, plecandu-se de la 10 cupluri, in prima
generatie au rezultat 813 indivizi, in cea de-a doua au rezultat 5.470de
indivizi, iar in cea de-a treia generatie au rezultat 188.324 de indivizi.
Deci populatia a crescut in trei generatii de 9416,2 ori. In ceea ce
priveste rata sexelor, daca in prima generatie a fost usor
favorabila masculilor (50,44%), in urmatoarele doua
generatii a fost favorabila femelelor (50,43%, respectiv 50,07%).
In ceea ce priveste
populatia Suceava, situatia se prezinta astfel: in prima
generatie in medie per cuplu au aparut 69,6 indivizi, ceea ce
totalizeaza o descendenta de 696 de indivizi, in cea de-a doua
generatie s-au obtinut 3.792 de indivizi, iar in a treia
generatie totalul indivizilor a fost de 122.658. Prin urmare, in ansamblu,
prolificitatea a fost mult mai scazuta in aceasta populatie
comparativ cu populatia Botosani, numarul indivizilor crescand
in cele trei generatii de 6132,2 ori. In ceea ce priveste rata
sexelor, in generatia I-a si a III-a au dominat masculii (51,28%
si, respectiv 51% ), femelele fiind procentual egale cu masculii (rata
sexelor =1) in generatia a II-a (50,01% femele).
Mentionam ca, in cele trei
generatii, pe parcursul carora s-au efectuat observatiile,
desi numarul de indivizi a fost destul de mare, nu s-au
evidentiat mutatii.
Tabel nr.1 - Datele sintetice
privind comportarea celor doua populatii, in trei generatii
succesive de consangvinizare
|
Generatia
|
Timpul necesar aparitiei tuturor indivizilor
|
Numarul total de indivizi
|
Rata sexelor (♂/♀)
|
|
Populatia Botosani
|
|
|
|
|
I
|
6,9 zile
|
|
|
|
II
|
10 zile
|
|
|
|
III
|
9 zile
|
|
|
|
Populatia Suceava
|
|
|
|
|
I
|
6,4 zile
|
|
|
|
II
|
9,6 zile
|
|
|
|
III
|
6,8 zile
|
|
|
VI.2. Efecte ale tratamentului cu nicotina, in diferite
concentratii, aplicat indivizilor de Drosophila
melanogaster dintr-o populatie reversmutanta
In primul
experiment, in urma tratamentului cu nicotina 0,005%, au rezultat 1158 de
indivizi (587 ♀ si 571 ♂) cu o medie de 129 indivizi per
eprubeta. Durata aparitiei indivizilor adulti s-a prelungit,
comparativ cu martorul, cu doua zile. Sub aspect numeric, tot prin
comparatie cu martorul, in medie per eprubeta, au aparut mai
multi indivizi (129 indivizi fata de 100 la martor).
Analizand
frecventa aberatiilor, la varianta de tratament si la martor,
constatam ca in cazul tratamentului cu nicotina 0,005%
frecventa aberatiilor scade (3,98 fata de 13,9 la martor).
Prin urmare, ca o prima concluzie, putem aprecia ca nicotina in
concentratia mentionata stimuleaza prolificitatea,
prelungeste perioada de aparitie a adultilor si reduce
frecventa mutatiilor. Altfel spus, nicotina in aceasta
concentratie are efecte benefice.
In cazul
administrarii nicotinei in concentratia de 0,01%, constatam
ca, per total experiment au aparut 1470 indivizi, ceea ce
reprezinta o medie de 163 indivizi per eprubeta, in timp ce la martor
au aparut 111 indivizi per eprubeta. Durata perioadei aparitiei
adultilor s-a prelungit, fata de martor, tot cu doua zile
(7 in loc de 5). Sub aspectul aberatiilor, de aceasta data
constatam ca administrarea nicotinei in concentratia de 0,01% a
determinat o scadere drastica a procentului acestora (2,16%) atat
fata de concentratia anterioara, cat si fata
de martor (12,18%). Prin urmare, efectele benefice sesizate la prima
concentratie, de aceasta data s-au amplificat.
Administrarea
nicotinei in concentratia de 0,03% a evidentiat urmatoarele
aspecte: in ceea ce priveste numarul total de indivizi si cel
per eprubeta, constatam scaderi marcante fata de
situatia concentratiilor anterioare si fata de martor
(247 indivizi per total, 39 indivizi per eprubeta). Perioada de
aparitie a adultilor s-a extins la zece zile. Prin urmare, viteza de
succesiune a generatiilor se reduce.
Din punct de
vedere al frecventei aberatiilor, am constatat o crestere a
acestora (6,07%), comparativ cu cele inregistrate la concentratiile
anterioare (3,98% in cazul concentratiei de 0,005% si 2,16% in cazul
concentratiei de 0,1%), mentinandu-se, totusi, sub
frecventa inregistrata la martor (14,67%).
Toate
constatarile mentionate mai sus, sunt ilustate in tabelele si
graficele, pe care vi le redam in continuare:
Tabel nr.2 - Media numerica a indivizilor din probele care au fost
supuse tratamentului cu nicotina 0,005%
|
Total indivizi in experimentul
1
|
Total indivizi in experimentul
2
|
Total indivizi in experimentul
3
|
TOTAL
general
|
MEDIA
generala
|
|
Epr.I
|
Epr.II
|
Epr.III
|
Epr.I
|
Epr.II
|
Epr.III
|
Epr.I
|
Epr.II
|
Epr.III
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.3 - Media numerica si procentuala a indivizilor
din probele martor in cazul tratamentului cu nicotina 0,005%
|
Martor proba A
|
Martor proba B
|
Martor proba C
|
TOTAL GENERAL
|
MEDIA GENERALA
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.4 - Media indivizilor cu aberatii in probele martor si
in cele supuse tratamentului cu nicotina 0,005%
|
Media indivizilor cu
aberatii in probele martor
|
Media indivizilor cu
aberatii in cazul tratamentuli cu nicotina
|
|
Nr.
|
|
Nr.
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.5- Media numerica a indivizilor din probele care au fost
supuse tratamentului cu nicotina 0,01%
|
Total indivizi in experiment 1
|
Total indivizi in experiment 2
|
Total indivizi in experiment 3
|
TOTAL
general
|
MEDIA
generala
|
|
Epr.I
|
Epr.II
|
Epr.III
|
Epr.I
|
Epr.II
|
Epr.III
|
Epr.I
|
Epr.II
|
Epr.III
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.6 - Media numerica si procentuala a indivizilor
din probele martor in cazul tratamentului cu nicotina 0,01%
|
Martor proba A
|
Martor proba B
|
Martor proba C
|
TOTAL GENERAL
|
MEDIA GENERALA
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.7 - Media indivizilor cu aberatii in probele martor si
in cele supuse tratamentului cu nicotina 0,01%
|
Media indivizilor cu
aberatii in probele martor
|
Media indivizilor cu
aberatii in cazul tratamentului cu nicotina
|
|
Nr.
|
|
Nr.
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.8 - Media numerica a indivizilor din probele care au fost
supuse tratamentului cu nicotina 0,03%
|
Total indivizi in experiment 1
|
Total indivizi in experiment 2
|
Total indivizi in experiment 3
|
TOTAL
general
|
MEDIA
generala
|
|
Epr.I
|
Epr.II
|
Epr.III
|
Epr.I
|
Epr.II
|
Epr.III
|
Epr.I
|
Epr.II
|
Epr.III
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.9 - Media numerica si procentuala a indivizilor
din probele martor in cazul tratamentului cu nicotina 0,01%
|
Martor proba A
|
Martor proba B
|
Martor proba C
|
TOTAL GENERAL
|
MEDIA GENERALA
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabel nr.10 - Media indivizilor cu
aberatii in probele martor si in cele supuse tratamentului cu
nicotina 0,03%
|
Media indivizilor cu
aberatii in probele martor
|
Media indivizilor cu
aberatii in cazul tratamentuli cu nicotina
|
|
Nr.
|
|
Nr.
|
|
|
|
|
|
|
|
|
|
|
|
Grafic nr. 1 Numarul total de indivizi aparuti in urma
tratamentului cu nicotina (concentratii diferite), comparativ cu
martorul
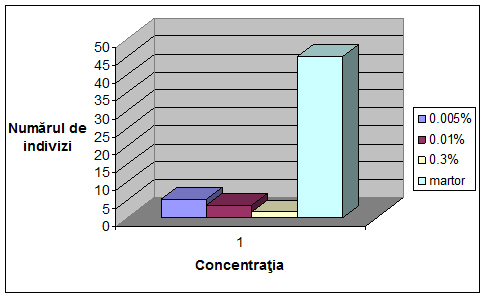
Grafic nr. 2 Numarul indivizilor cu
aberatii aparuti in cazul tratamentului cu nicotina,
si in proba martor
VI.3. Dinamica fondului de gene in populatii experimentale de
Drosophila melanogaster sub presiune selectiva
Tabel nr. 11 - Dinamica genotipului/zile de prelevare, in
populatii de Drosophila melanogaster ( Experiment I, prima generatie)
Genotipul
|
Numarul de indivizi pe
zile de prelevare (F1)
|
|
Ziua I
|
Ziua II
|
Ziua III
|
Ziua IV
|
Ziua V
|
Total genotip
|
Total experiment
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
Oregon (70)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Brown
Bw (5)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vestigial
Vg(240)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
White
W(111)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Black
B(46)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
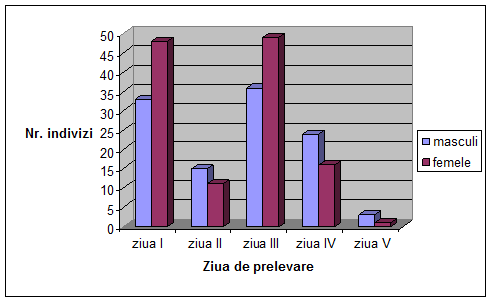
Grafic nr. 3 - Variatia nr. de indivizi Oregon/sexe/zile de prelevare
(Experiment I, prima generatie)
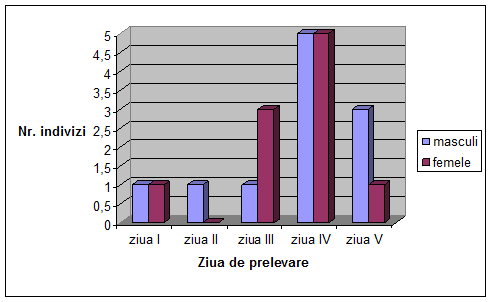
Grafic nr. 4 - Variatia nr. de indivizi Brown/sexe/zile de prelevare
(Experiment I, prima generatie)
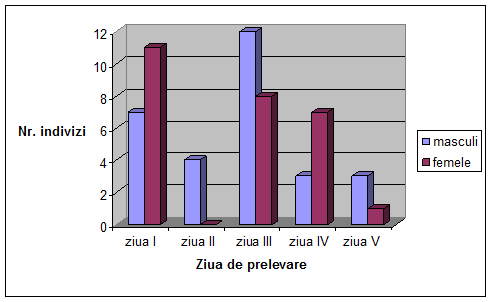
Grafic nr. 5 - Variatia nr. de indivizi Vestigial/sexe/zile de
prelevare (Experiment I, prima generatie)
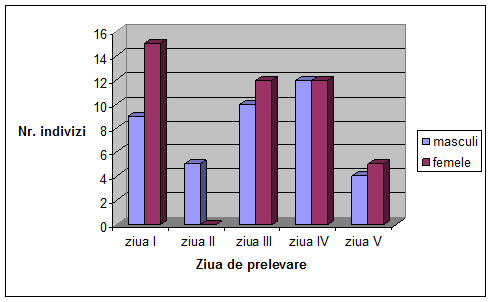
Grafic nr. 6 - Variatia nr. de indivizi White/sexe/zile de prelevare
(Experiment I, prima generatie)
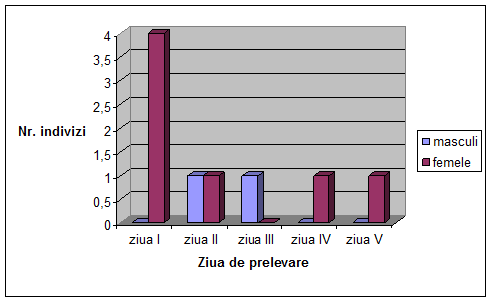
Grafic nr. 7 - Variatia nr. de indivizi Black/sexe/zile de prelevare
(Experiment I, prima generatie)
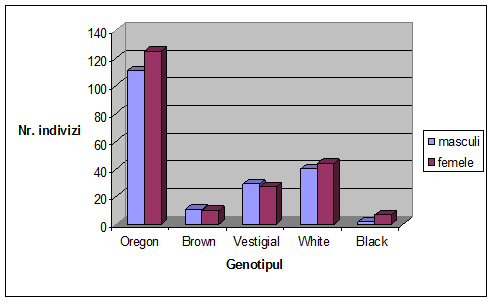
Grafic nr. 8 - Variatia nr. de indivizi /sexe /genotip (Experiment I,
prima generatie)
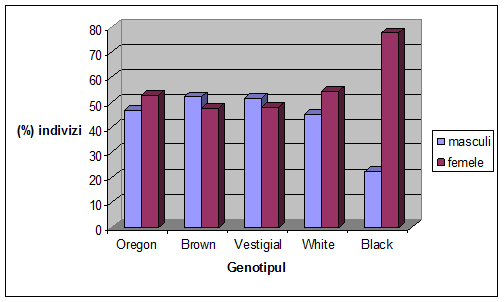
Grafic nr. 9 - Variatia procentuala a nr. de indivizi /sexe
/genotip (Experiment I, prima generatie)
Tabel nr. 12 - Dinamica genotipului/zile de prelevare, in
populatii de Drosophila melanogaster ( Experiment I, a II-a generatie)
Genotipul
|
Numarul de indivizi pe
zile de prelevare (F2)
|
|
Ziua I
|
Ziua II
|
Ziua III
|
Ziua IV
|
Ziua V
|
Total genotip
|
Total experiment
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
Oregon (70)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Brown
Bw (5)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vestigial
Vg(240)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
White
W(111)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Black
B(46)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
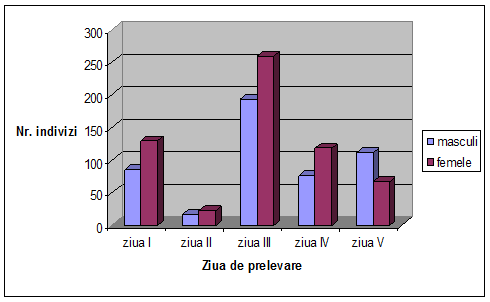
Grafic nr. 10 - Variatia nr. de indivizi Oregon/sexe/zile de
prelevare (Experiment I, generatia a II-a)
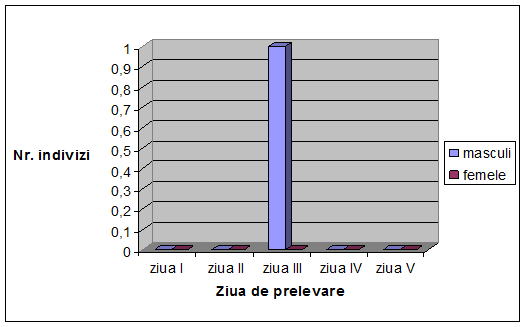
Grafic nr. 11 - Variatia nr. de indivizi Brown/sexe/zile de prelevare
(Experiment I, generatia a II-a)
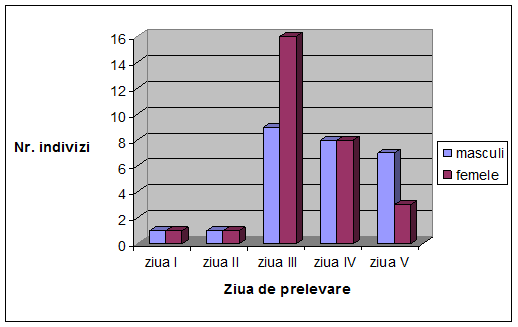
Grafic nr. 12- Variatia nr. de indivizi Vestigial/sexe/zile de
prelevare (Experiment I, generatia a II-a)
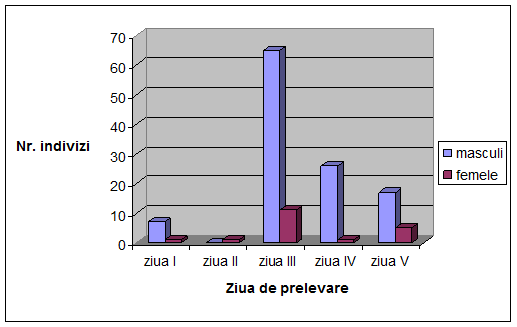
Grafic nr. 13 - Variatia nr. de indivizi White/sexe/zile de prelevare
(Experiment I, generatia a II-a)
Grafic nr. 14 - Variatia nr. de indivizi Black/sexe/zile de prelevare
(Experiment I, generatia a II-a)
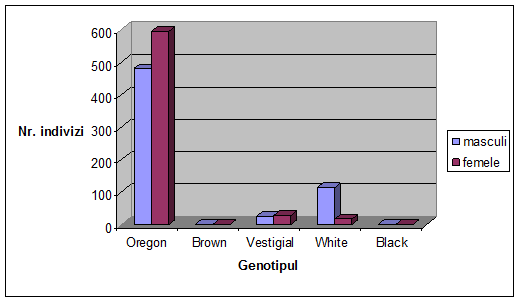
Grafic nr. 15 - Variatia nr. de indivizi /sexe /genotip (Experiment
I, generatia a II-a)
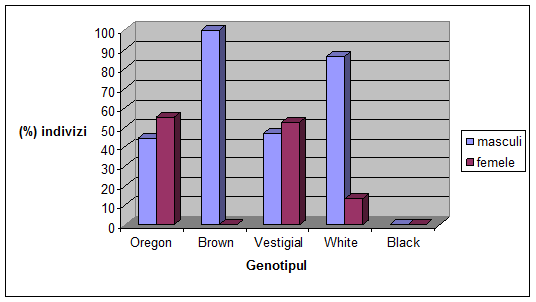
Grafic nr. 16 - Variatia procentuala a nr. de indivizi /sexe
/genotip (Experiment I, generatia a II-a)
Tabel nr. 13 - Dinamica genotipului/zile de prelevare, in
populatii de Drosophila melanogaster ( Experiment II, prima generatie)
Genotipul
|
Numarul de indivizi pe
zile de prelevare (F1)
|
|
Ziua I
|
Ziua II
|
Ziua III
|
Ziua IV
|
Ziua V
|
Total genotip
|
Total experiment
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
Oregon (70)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Curly
Cy(10)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vestigial
Vg(240)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
White
W(111)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Black
B(46)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
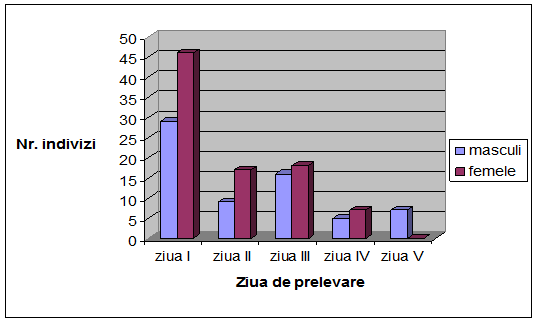
Grafic nr. 17 - Variatia nr. de indivizi Oregon/sexe/zile de
prelevare (Experiment II, prima generatie)
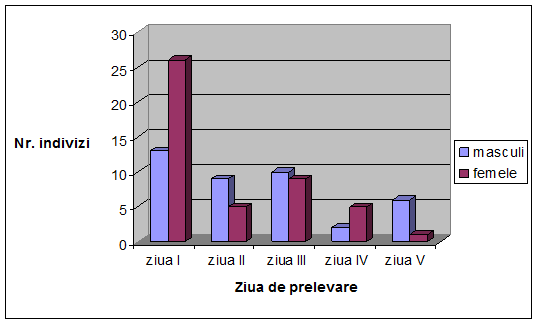
Grafic nr. 18 - Variatia nr. de indivizi Curly/sexe/zile de prelevare
(Experiment II, prima generatie)
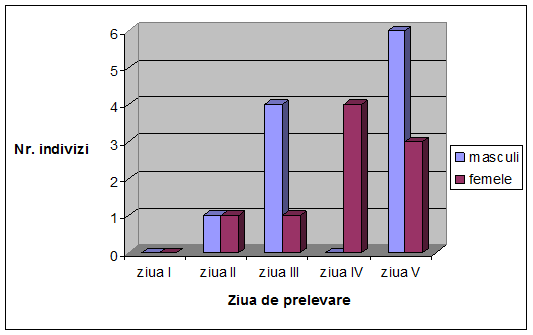
Grafic nr. 19 - Variatia nr. de indivizi Vestigial/sexe/zile de
prelevare (Experiment II, prima generatie)
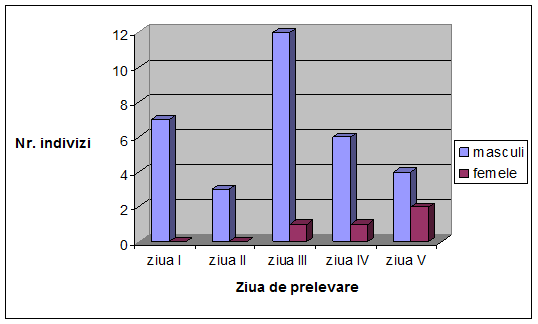
Grafic nr. 20 - Variatia nr. de indivizi White/sexe/zile de prelevare
(Experiment II, prima generatie)
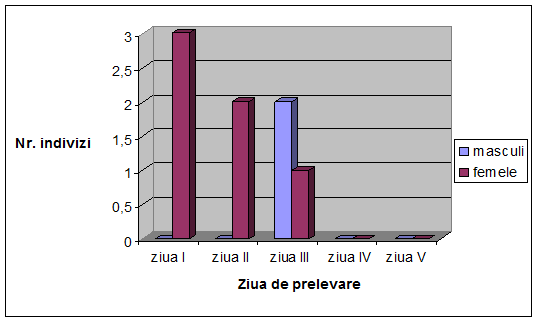
Grafic nr. 21- Variatia nr. de indivizi Black/sexe/zile de prelevare
(Experiment II, prima generatie)
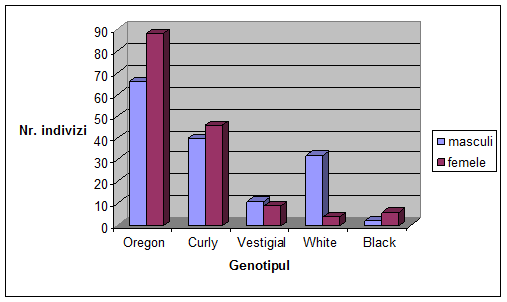
Grafic nr. 22- Variatia nr. de indivizi /sexe/genotip (Experiment II,
prima generatie)
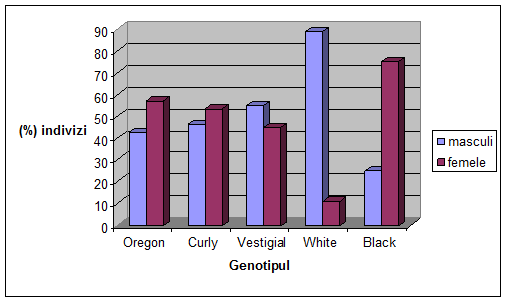
Grafic nr. 23 - Variatia procentuala a nr. de indivizi
/sexe/genotip (Experiment II, prima generatie)
Tabel nr. 14 - Dinamica
genotipului/zile de prelevare, in populatii de Drosophila melanogaster
(Experiment II, a II-a generatie)
Genotipul
|
Numarul de indivizi pe zile de prelevare (F1)
|
|
Ziua I
|
Ziua II
|
Ziua III
|
Ziua IV
|
Ziua V
|
Total genotip
|
Total experiment
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
Nr.
|
|
|
Oregon (70)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Curly
Cy (10)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vestigial
Vg(240)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
White
W(111)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Black
B(46)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
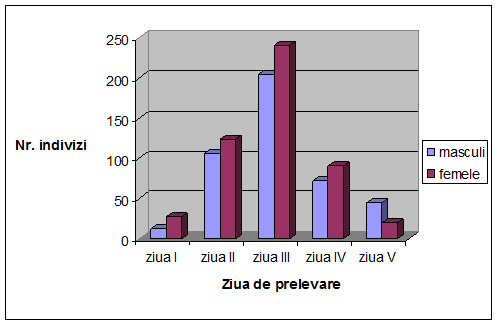
Grafic nr. 24- Variatia nr. de indivizi Oregon/sexe/zile de prelevare
(Experiment II, generatia a II-a)
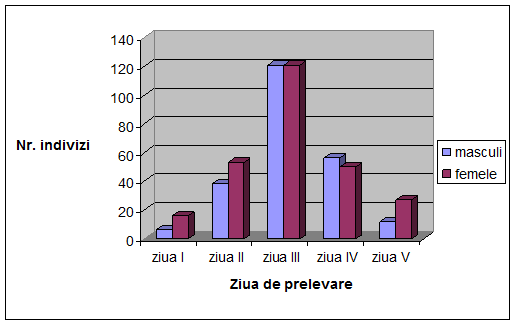
Grafic nr. 25 - Variatia nr. de indivizi Curly/sexe/zile de prelevare
(Experiment II, generatia a II-a)
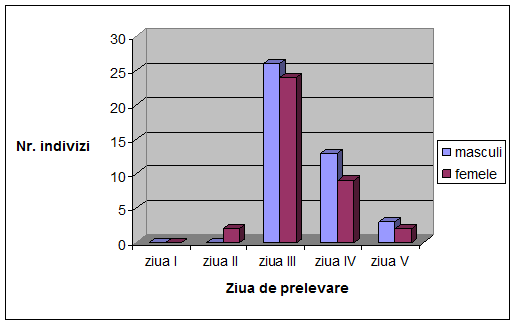
Grafic nr. 26 - Variatia nr. de indivizi Vestigial/sexe/zile de
prelevare (Experiment II, generatia a II-a)
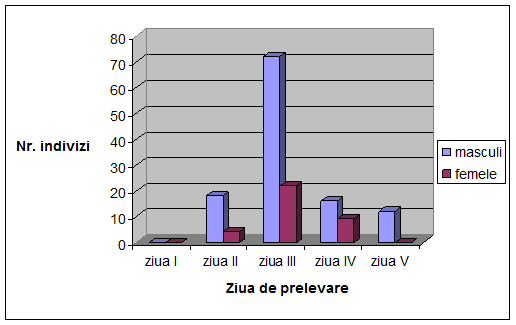
Grafic nr. 27 - Variatia nr. de indivizi White/sexe/zile de prelevare
(Experiment II, generatia a II-a)
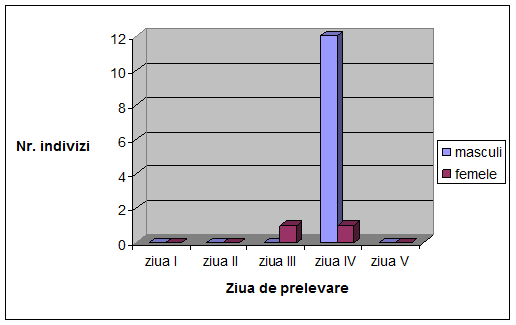
Grafic nr. 28 - Variatia nr. de indivizi Black/sexe/zile de prelevare
(Experiment II, generatia a II-a)
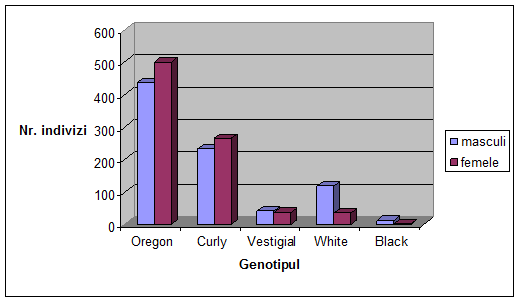
Grafic nr. 29 - Variatia numarului de indivizi /sexe/genotip
(Experiment II, prima generatie)
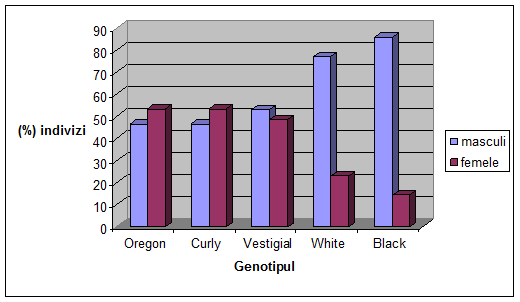
Grafic nr. 30 - Variatia procentuala a numarului de
indivizi /sexe /genotip (Experiment II, generatia a II-a)
In cadrul celui de-al II-lea
experiment, in generatia a II-a s-au obtinut indivizi Curly dublu
mutanti, ca a doua mutatie aparand culoarea alba a ochilor.
Astfel din 500 indivizi Curly obtinuti, 59 prezentau cea de-a doua
mutatie - ochi de culoare alba.
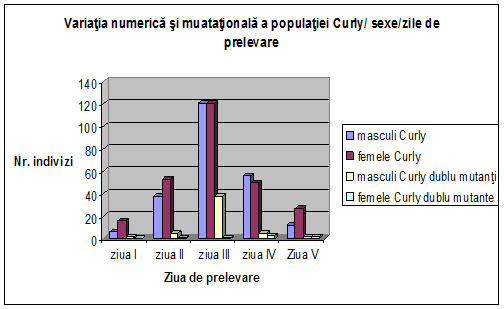
Grafic nr. 31 - Variatia
numerica a mutatiilor in a populatia Curly/sexe/zile de
prelevare
CONCLUZII
Pe baza investigatiilor efectuate
in prezenta cercetare, am ajuns la urmatoarele concluzii:
Efecte ale consangvinizarii in doua populatii locale de Drosophila melanogaster
Rata
natalitatii, sub impactul consangvinizarii, s-a mentinut
mult mai ridicata in cadrul populatiei Botosani, comparativ cu
populatia Suceava.
Tinand
cont ca succesiunea generatiilor este mai rapida la
populatia Suceava, putem concluziona ca, in timp mai indelungat,
numarul descendentilor per cuplu poate ajunge mai mare decat
populatia Botosani.
Rata sexelor
a fost echilibrata, in ambele populatii, cu o usoara
dominanta a femelelor in populatia Botosani si a
masculilor in populatia Suceava.
In nici una
dintre populatii nu s-au evidentiat mutante.
Efecte ale tratamentului cu nicotina, in diferite concentratii,
aplicat indivizilor de Drosophila
melanogaster dintr-o populatie reversmutanta
Nicotina, in
concentratiile de 0,005% si 0,01%, are efecte pozitive asupra
prolificitatii indivizilor de Drosophila
melanogaster.
Toate cele
trei concentratii, administrate, au prelungit perioada de aparitie a
indivizilor, pe mediul de cultura, in conditii de laborator. Cea mai
lunga perioada (10 zile) s-a inregistrat sub efectul
concentratiei de 0,03%.
Putem
aprecia, totusi, ca desi nicotina in concentratii mici (sub
0,015%) are efecte benefice asupra prolificitatii, avand in vedere
ca prelungeste cilclul de dezvoltare al indivizilor, intr-un interval
de timp mai lung numarul total al indivizilor poate fi mai mic decat la
martor.
Nicotina a
determinat o reducere a frecventei aberatiilor, mai ales in
concentratia de 0,01% si apoi in cea de 0,005%.
Dinamica fondului de gene in populatie
experimentala de Drosophila melanogaster sub presiune selectiva
Rata natalitatii, sub
presiunea selectiei naturale, s-a mentinut ridicata pe parcursul
ambelor experimente in cazul tipului salbatic (Oregon), comparativ cu cu populatiile celorlalte mutante.
In cadrul celui de-al doilea
experiment, in generatia a doua s-a observat ca populatia Curly
prezinta o mare capacitate mutationala.
BIBLIOGRAFIE:
ABRAHAMSON, S., LEWIS E. B., 1971. The detection of mutations in Drosophila
melanogaster, in: Chemical mutagens: Principles and methods for their detection,
A. Hollander, ed., vol. 2, Plenum Press, Oxford, pp. 461-487.
ASHBURNER M., THOMPSON J. N., 1978 : The Genetics and Biology of
Drosophila, vol. 2A, ASHBURNER M., Wright T. R. F., eds., Academic Press, London.
ASHBURNER M., Wright, T. R. F., 1978 : The Genetics and Biology of Drosophila, vol. 2B, Academic Press,
London, New York, San Francisco, 500-504
BARA, I. I., 1996. Vademecum in genetica. Editura Corson, Iasi
BARA, I. I., 1973. Studiul asupra biologiei populatiilor unor specii apomictice
si sexuate inrudite. Teza de doctorat, Facultatea de Biologie,
Universitatea Bucuresti.
BARA, I. I., 1999. Genetica. Editura Corson, Iasi
BARA, I., ION, SURUGIU, C., IULIANA, CIMPEANU, M.
MIRELA, MANIU, MARILENA. GENETICA SI EVOLUTIONISM. Editura Corson, Iasi
2000.
BORROR, D.J., DE LONG D.M., 1964. An Introduction to the Study of Insects. Holt, Rinehart and
Winston, Inc., New York, Chicago, San Francisco, Toronto, London.
BOTNARIUC, N., 1976. Conceptia
si metoda sistemica in biologia generala. Editura Academiei,
Bucuresti.
BOTNARIUC, N., 1992. Evolutionismul
in impas Editura Academiei, Bucuresti.
BOTNARIUC, N., 1999. Evolutia
sistemelor biologice supraindividuale. Editura Universitatii
Bucuresti.
BRIGGS, D., WALTERS, S. M., 1969. Plant Variation and Evolution. McGraw-Hill, New York, Toronto.
CARMEN, S., 1996, Mutageneza
chimica, Ed. Genesis, Cluj-Napoca.
CIOBOTARI, GH., BARA, I.I., 200. Efecte ale tratamentului cu nicotina,
in diferite concentratii, aplicat indivizilor de vDrosophila melanogaster,
intr+o populatie revers-mutanta.In: Genetica si evolutionism (editata de Bara si colab., 2000), Editura
CORSON-Iasi, 79-84
CIRLAN, M., CREANGA, ST., 2001. Evolutia determinismului genetic al
sexelor. Editura Sedcom Libris, Iasi.
COOK, L.M., 1976. Population
Genetics. Chapman and Hall, A Halsted Press Book London, John Wiley &
Sons, Inc., New York.
CRACIUN, T., I., TOMOZEI, N., COLES, A., NASTA, Genetica, Editura Didactica si Pedagogica
Bucuresti - 1978.
CREED, E. R., 1971. Ecological
Genetics and Evolution. Essays in Honour of E. B. Ford. Blackwell, Oxford.
CROW, J. F., KIMURA, M., 1970. An Introduction to Population Genetic Theory. Harper and Row, New
York.
DARWIN, CH., WALLACE, A. R., 1958. Evolution by natural selection. Cambridge University Press.
DOBZHANSKY, T., 1970. Genetics of
the evolutionary process. Columbia University Press, New York.
DUBININ, N. P., 1985, Genetica,
Chisinau, Stiinta;
DUBININ, N. P., 1977, Miscarea
eterna, Editura Politica, Bucuresti;
ENESCU V., 1985, Genetica
Ecologica, Ed. Ceres, Bucuresti;
FALCONER, D. S., 1960. Introduction
to quantitative Genetics. Oliver and Boyd, Edinburgh.
FORD, E. B., 1975. Ecological
Genetics, 4th Edition. Chapman and Hall, London.
GLASS, BENTLEY, SACKS, M. S., JAHN, E. F., HESS, C., 1952. Genetic Drift in a Religious Isolate: An
Analysis of the Causes of Variation in Blood Group and Other Gene Frequencies in a Small Population.
American Naturalist, 86, 145-160.
HENLE, K., 1991. Some
reflections on evolutionary theories, with a classification of fitness.
Acta Biotheoretica, 39, 91-106.
KIMURA, M., OHTA, T., 1971. Theoretical Aspects of Population Genetics. Princeton University
Press, Princeton, New York.
KING, J. L., JUKES, T. H., 1969. Non-Darwian evolution. Science, 164, 788-798.
KREBS, CH. J., 1972. Ecology. The
Experimental analysis of distribution and abundance. Harper & Row
Publishers, New York, Evanston, San Francisco, London.
METTLER, L.E., GREGG, T.G., 1974, Genetica populatiilor si evolutia, Editura
Stiintifica Bucuresti;
MOVILEANU V., POPA N., 1996, Genetica. Lucrari practice, Editura Stiinta,
Chisinau;
PALII, A., 1998, Genetica,
Ed. Musseum.
RACOVITA, E., 1929. Evolutia si problemele ei. Editura eugenica si
biopolitica a Astrei, IV, Cluj, 183 pg.
RAICU P., NACHTIGAL M., 1969, Citogenetica. Principii si metode, Editura Academiei,
Bucuresti;
RAICU, P., 1980. Genetica.
Editura Didactica si Pedagogica, Bucuresti.
SABASANU, IONELA. 2000. Complementaritatea intergenica in cazul
incrucisarilor reciproce sepia x vermilion si Müller 5 x
vermilion, la Drosophila melanogaster. Lucrare de dizertatie,
Universitatea "Al. I. Cuza" Iasi
SIMPSON, G. G., 1951. The meaning
of evolution. New American Library, New York.
SLATKIN, M., 1985. Gene flow in
natural populations. Annual Reviw af Ecology and Systematics, 16, 393 -
430.
STRICKBERGER, M.W., 1962. Experiments in Genetics with Drosophila. John Wiley & Sons,
Inc., New York, London, Sydney.
STUGREN, B., 1969. Evolutionismul
in secolul 20. Certitudini. Divergente. Idealuri. Editura
Politica, Bucuresti.
TOMA, N., GAVRILA, L., 2000. Ereditatea extranucleara. Editura Universitatii din
Bucuresti.
TUDOSE, I. GH., 1992 - 1993. Genetica
vol I - II. Editura Univ. "Al. I. Cuza" Iasi.
TUDOSE I.GH.,
BRANDSCH R., 1969, Modificari in
desfasurarea ciclului vital la Drosophila melanogaster, sub
influenta efedrinei si influenta acestei substante asupra
cromosomilor uriasi. Cercetari de Genetica, Editura
Didactica si Pedagogica, Bucuresti, 404-407.

