SISTEMUL
COMPLEMENT (COMPLEMENTUL SERIC)
Definitie
« Complementul
seric reprezinta un grup de proteine si glicoproteine serice care se
activeaza secvential, interactioneaza unele cu altele, cu
anticorpi din complexele imune si cu receptorii de suprafata ai
celulelor fagocitare producand multe semnale chimice si un raspuns
distinctiv cu rol de protectie nespecifica fata de
diversi patogeni ».
Principalele roluri ale complementului seric sunt:
a)
liza microorganismelor patogene;
b)
opsonizarea bacteriilor;
c)
chemotactism pentru celule;
d)
clearance-ul complexelor imune.
Pana
in prezent se cunosc trei cai de
activare a complementului seric:
calea clasica - ce necesita
prezenta complexelor Ag - Ac.
Aceasta este calea imunitatii specifice.
calea alternativa - ce este
actionata prin toxine;
calea lectinica (prin lectine
legate pe suprafata microorganismelor), cum este MBP (mannose binding
protein) - o apoproteina serica, ce leaga manoza de la componentii
glucidici din invelisul bacterian sau viral.
1.
Calea clasica de activare a complementului
Activarea complementului seric pe calea clasica se realizeaza
dupa fixarea anticorpilor pe Ag.Cele
11 componente care participa la activarea complementului pe calea
clasica sunt organizate in trei
unitati functionale:
unitatea de
recunoastere - C1a (C1q, C1r, C1s);
unitatea de activare (C4 C2 C3);
unitatea de atac (C5b6789);
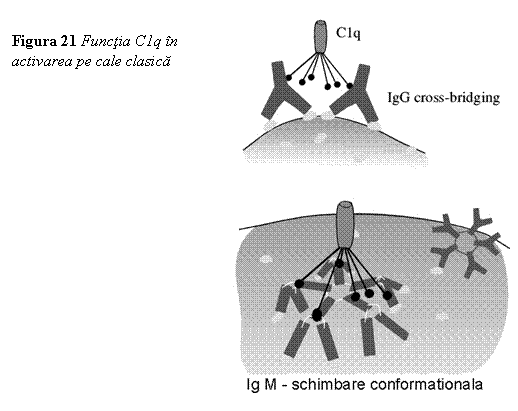
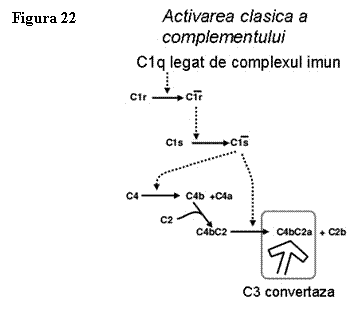
Activarea
complementului pe cale clasica (fig. 21, 22) incepe prin legarea complexelor
Ag-Ac de membrana celulara, ceea ce duce la modificari in structura conformationala a moleculei de
Ig, modificari care sunt recunoscute de C1q in prima faza, iar apoi si de C1r si C1s ceea ce
duce la formarea C1qrs care se mai numeste si C1 - esteraza si care are
activitate enzimatica in prezenta ionilor de calciu.
C1-esteraza cliveaza C4 in doua fragmente
inegale C4a si C4b, dar si pe C2 in C2a si C2b.
C4b se leaga pe membrana celulara langa complexul Ag-Ac.
C2a, in prezenta ionilor de Mg, se combina cu C4b
de pe membrana celulara si formeaza C4bC2a, care se numeste C3 -
convertaza, enzima ce scindeaza C3 (componentul central al complement
seric) in C3a (solubil) - cu rol in
declansarea inflamatiei si C3b
- care se leaga pe de-o parte de suprafata (cu rol in
aderenta si opsonizare, in inactivare) si pe de alta
parte de C4bC2a, cu care
formeaza C4bC2aC3b numit C5 - convertaza.
Acesta
scindeaza C5 in C5a si C5b.
C5b leaga C6
si C7 si formeaza C5b67 cu
rol chemotactic pentru neutrofile si monocite. Acest complex se leaga
de membrana celulara, si de el se leaga apoi C8 si C9 si se formeaza
complexul C5b6789 numit complex
membranar de atac (MAC).
Complexul
membranar de atac este imaginat ca un canal transmembranar foarte
permeabil pentru apa si electroliti care permite traversarea
apei si electrolitilor cu modificarea gradientului protonic (H+). Ca
urmare are loc lizarea celulei, adica un soc osmotic.
Activarea
complementului seric pe calea clasica este controlata de doua proteine serice:
inactivatorul lui C1q care blocheaza
activitatea enzimatica a C1-esterazei;
inactivatorul lui C3b care scindeaza C3b in
2 componente inactive: C3c si C3d.
2. Activarea complementului pe calea
alternativa
Activarea
pe cale alternativa a complementului este realizata de:
polizaharide (din bacterii, fungi);
lipopolizaharide (endotoxinele bacteriilor Gram-negative);
un factor din veninul cobrei (VCF);
IgA agregata;
de complexe Ag-Anticorp care nu activeaza calea
clasica.
Cele doua cai de activare au in comun
componentul C3b care se leaga pe membrana celulara de C4bC2a -
denumit C5-convertaza caii clasice.
C3b, sub influenta ionilor de Mg2+ si a unor activatori, se combina cu factorul B (care
este echivalentul lui C2) si formeaza
C3bB, care in prezenta factorului D (o proteaza plasmatica activata de Mg2+), se
poate transforma in C3bBb (dupa
ce factorul B a fost scindat in Ba si Bb). C3bBb, numit si C3-convertaza
caii alternative, are o activitate enzimatica importanta: descompune moleculele de C3.
Clivarea C3 este foarte importanta in procesele de
aparare anti-microbiana.
Un
numar mare de microorganisme pot activa C3Bb si pot produce cantitati mari de produsi
de clivare ai C3, cu producerea mai
multor molecule de C3b si
fixarea lor pe suprafata microorganismelor, antrenand activarea
urmatorului component - C5,
clivarea lui in C5a si C5b
(fixat pe membrana).
Urmeaza
apoi activarea celorlalte componente C6,C7,C8,C9
cu formarea MAC (la fel ca si in cazul caii clasice).
Importanta caii alternative consta in
faptul ca prin aceasta se realizeaza activarea complementului seric
prin produsele bacteriene, inaintea sintezei de Ig specifice.
3. Componentele reglatorii ale
complementului
Fragmentele
cheie ale activarii complementului sunt rapid inactivate daca ele
raman in stare libera.
Proteinele
reglatoare ale activarii complementului sunt :
C1 - inhibitor care
se leaga covalent de C1rC1s ceea ce va duce la disocierea C1rC1s de C1q
care ramane legata de anticorp fapt ce duce la limitarea timpului in
care C1qrs poate cliva C4. Absenta C1 - INH duce la aparitia
angioedemului ereditar;
La nivelul caii
clasice C4aC2b este un compus foarte labil fiind inactivat spontan prin
desprinderea componentei C2b;
C4BP (C4-bingding
protein) se leaga de C4b ceea ce permite :
clivarea C4b de catre factorul I cu formare de C4c
si C4d - care ramane atasat de membrana bacteriana si
impiedica legarea C2b de aceasta;
favorizeaza disocierea C2b din C4b2b;
DAF (Decay Accelerating Factor) amplifica disocierea C2b
din C4b2b;
Factorul S (vitronectina) impiedica formarea si
insertia MAC.
4. Evaluarea complementului
Metodele de determinare sunt:
Cantitative - imunodifuzia radiala simpla Mancini
(IDRS), turbimetrica,
nefelometrica, imunoenzimatica,
radioimunologica.
Semicantitative - imunoelectroforeza in "racheta"
Analiza cantitativa
Proteinele
sistemului complement pentru investigarea caii clasice sunt:
C1q
- 10-25 mg%;
C1r
- 5mg%;
C1s
- 5mg%;
C4
- 20-50mg%;
C2
- 2,5mg%.
Proteinele
sistemului complement pentru investigarea caii alternative sunt:
C3
- 55-120mg%;
C3a
- 120mg%:
Factorul
D;
Factorul
B.
Proteinele
sistemului complement pentru investigarea secventei terminale sunt:
C5,
C5a - 7 mg%;
C6
- 6 mg%;
C7
- 5 mg%;
C8
- 5,5 mg%;
C9
- 6 mg%.
Determinarea activitatii hemolitice a serului
Dozarea activitatii hemolitice a
complementului
Complement
hemolitic total: CH50 = 45-55 u/ml = calea clasica;.
Complement
hemolitic: APH50 sau AP50= 15-20u /ml calea alternativa.
Dozarea componentilor complementului
Dozarea
proteinelor inhibitorii.
5.
Importanta sistemului complement in imunopatologie
Deficienta
uneia sau mai multor componente ale sistemului complement duce la aparitia
pe plan clinic a:
cresterii
susceptibilitatii la infectii;
bolilor
de colagen;
angioedemului.
Astfel deficitul
factorilor C1, C2, C4, C3 determina o susceptibilitate crescuta
fata de infectiile cu bacterii incapsulate (Pneumococ,
Haemophilus I. etc) datorita faptului ca nu se mai produce
opsonizarea acestor bacterii care este esentiala pentru
fagocitoza.
Deficitul de C5-9
(MAC) duce la infectii sistemice cu Neisseria.
Deficientele
componentelor C1, C2, C4, C3 determina si aparitia unor boli de
colagen cum ar fi: lupusul eritematos sistemic, dermatomiozita, dar si a
unor vasculite, glomeulonefrite membranoproliferative, purpura
anafilactoida.
Deficitul de C1q:
se
transmite autosomal recesiv;
poate
fi de doua tipuri:
absenta
completa a componentei C1q;
C1q
prezenta, dar nefunctionala.
Clinic apare:
sindrom
lupus-like cu: febra, rush, artrita, glomerulonefrita;
cresterea
susceptibilitatii la infectii.
Deficitul de
C1q/C1r
determina
aparitia unui sindrom lupus-like.
Deficitul C4:
poate
fi un deficit complet a componentei C4 care determina:
imposibilitatea
activarii complementului pe cale directa;
pastrarea posibilitatii activarii pe
cale alternativa, dar mai redusa.
clinic
se manifesta ca un sindrom lupus-like sau printr-o crestere a
susceptibilitatii la infectii;
poate
exista deficienta unui izotip C4a sau C4b;
deficitul
de C4a determina incapacitatea de procesare eficienta a complexelor
imune ce contin proteine proprii, ceea ce duce la cresterea riscului
pentru aparitia lupusului eritematos diseminat;
deficitul
de C4b duce la incapacitatea de activare a complementului pe cale directa
in prezenta unor suprafete bacteriene bogate in carbohidrati,
ceea ce determina o crestere a predispozitiei spre septicemii
si meningite.
Deficitul de C2:
este
rar 1:10.000 indivizi;
clinic apare:
crestere
a susceptibilitatii la infectii mai ales cu bacterii piogene
incapsulate (pneumococ, H.Influentzae, meningococ);
cresterea
prevalentei bolilor reumatice.
Deficitul de C3:
se
transmite autosomal recesiv;
determina
scaderea marcata a capacitati de opsonizare si a
activarii MAC;
clinic apare:
cresterea
susceptibilitatii la infectii cu bacterii piogene incapsulate
(pneumococ, meningococ);
afectiuni
reumatice: artralgii, rush cutanat, sindrom lupus-like, glomerulonefrita
membranoproliferativa.
Deficitul de C5:
transmitere
autosomal recesiva;
determina
infectii sistemice cu Neisseria si mai rar lupus eritematos
diseminat.
Deficitul de C6:
transmitere
autosomal recesiva;
manifestari
clinice identice cu deficitul de C5.
Deficitul de C7:
transmitere
autosomal recesiva;
manifestari clinice
de tip lupus eritematos sistemic, artrita reumatoida, sclerodermie.
Deficitul de C8:
transmitere
autosomal recesiva;
clinic apar manifestari similare cu C8.
Deficitul C9:
transmitere
autosomal recesiva;
susceptibilitate
la infectii sistemice cu Neisseria.
Deficitul
factorului I:
determina
lipsa de inactivare a C3 - convertaza ceea ce duce la o clivare spontana
si continua a complementului cu o scadere marcata a C3 in
ser;
clinic apar manifestari similare cu deficitul de C3.
Deficitul
factorului H:
foarte
rar;
clinic apare o crestere a susceptibilitatii
la infectii, afectiuni reumatice sau pacientii pot fi
asimptomatici.
Deficitul
properdinei:
transmitere
X-linkata;
determina
deficit de C3 - convertaza caii alternative care clinic se manifesta cu aparitia infectiilor
meningococice sistemice.
Deficitul de
C1-INH.
transmitere
autosomal dominanta;
clinic apar episoade recurente de edem subcutanat sau
submucos;
localizarea
poare fi la nivel:
 cutanat: extremitati, fata, zone
genitale. Zonele afectate sunt edematiate dar nu prezinta prurit sau
eritem;
cutanat: extremitati, fata, zone
genitale. Zonele afectate sunt edematiate dar nu prezinta prurit sau
eritem;
gastrointestinal, cu aparitia anorexiei, varsaturilor,
durerilor abdominale, a diareei. Imagistic se deceleaza edemul mucoasei
intestinale cu ingustarea sau obstructia lumenuluui intestinal;
respirator, la nivelul mucoasei bucale, linguale, orofaringiene,
laringiene.
DE RETINUT:
nu este afectata niciodata mucoasa respiratorie sub laringe astfel
ca traheostomia este totdeauna eficienta;